




Abstract
The ability of cannabinoids to modulate both inflammatory and degenerative neuronal damage prompted investigations on the potential benefits of such compounds in multiple sclerosis (MS) and in animal models of this disorder. Here we measured endocannabinoid levels, metabolism and binding, and physiological activities in 26 patients with MS (17 females, aged 19–43 years), 25 healthy controls and in mice with experimental autoimmune encephalomyelitis (EAE), a preclinical model of MS. Our results show that MS and EAE are associated with significant alterations of the endocannabinoid system. We found that anandamide (AEA), but not 2-arachidonoylglycerol (2-AG), was increased in the CSF of relapsing MS patients. AEA concentrations were also higher in peripheral lymphocytes of these patients, an effect associated with increased synthesis and reduced degradation of this endocannabinoid. Increased synthesis, reduced degradation, and increased levels of AEA were also detected in the brains of EAE mice in the acute phase of the disease, possibly accounting for its anti-excitotoxic action in this disorder. Accordingly, neurophysiological recordings from single neurons confirmed that excitatory transmission in EAE slices is inhibited by CB1 receptor activation, while inhibitory transmission is not. Our study suggests that targeting the endocannabinoid system might be useful for the treatment of MS.
Introduction
Immune-mediated attack of oligodendrocytes and myelin sheaths is a primary pathological event in multiple sclerosis (MS). Neurodegenerative damage, however, occurs early in this disorder, paralleling inflammation (Bjartmar et al., 2001; Peterson et al., 2001; De Stefano et al., 2002; Kuhlmann et al., 2002; Filippi et al., 2003; Filippi and Rocca, 2005). In recent years, the ability of cannabinoids to modulate both inflammatory (Baker et al., 2003; Walter and Stella, 2004), and degenerative neuronal damage (Grundy et al., 2001; Iversen, 2003) prompted investigations of the potential benefits of these compounds in MS, also encouraged by early reports affirming reduced frequency of relapses (Consroe et al., 1997), and relief from spasticity, dystonia, tremor, ataxia, bladder dysfunction and pain in MS patients self-medicating with marijuana (Consroe, 1998; Fernandez-Ruiz et al., 2002; Pertwee, 2002; Zajicek et al., 2003).
In experimental animal models of MS, cannabinoid administration significantly attenuates the clinical and pathological features of the disease (Lyman et al., 1989; Wirguin et al., 1994; Arevalo-Martin et al., 2003; Croxford and Miller, 2003), an effect also achieved by pharmacological interventions aimed at enhancing the levels of the endogenous cannabinoid anandamide (AEA) (Mestre et al., 2005; Ortega-Gutierrez et al., 2005; Bari et al., 2006; Ligresti et al., 2006). Furthermore, mice lacking the CB1 receptor develop more severe neuronal damage following experimental autoimmune encephalomyelitis (EAE) induction, supporting a role for endocannabinoids in this inflammatory neurodegeneration (Pryce et al., 2003).
Whether MS or experimental models of MS perturb the endocannabinoid system (ECS) is however controversial. Recently, increased levels of AEA, but not of the other abundant endocannabinoid 2-arachidonoylglycerol (2-AG), have been reported in tissues from inflammatory lesions of patients with MS (Eljaschewitsch et al., 2006). Another study, however, showed that the expression of the enzyme involved in AEA degradation was upregulated in MS lesions, indicating that the level of this endocannabinoid might be reduced in MS patients (Benito et al., 2007).
Furthermore, it has been reported that production of both AEA and 2-AG was normal in mice with asymptomatic to mild EAE (Shohami and Mechoulam, 2006; Witting et al., 2006), while both endocannabinoids were elevated in the chronic relapsing mouse model of MS (Baker et al., 2001). Finally, both AEA and 2-AG have been found to be down-regulated in the brains of rats with EAE (Cabranes et al., 2005). Decreased expression of CB1 receptors has also been reported in this rat model of MS (Berrendero et al., 2001), while the functionality of the same receptors seem to remain unchanged in mice with asymptomatic to mild EAE (Pertwee, 2002; Witting et al., 2006).
Thus, to increase our understanding of possible changes in ECS in MS, we measured endocannabinoid levels, metabolism and binding, and physiological activities in patients with MS and in mice with EAE. Our results indicate a profound rearrangement of the ECS involving AEA and the sensitivity of striatal cannabinoid CB1 receptors.
Subjects and Methods
The study was approved by the Ethics Committee of the University of Rome Tor Vergata.
MS patients
We collected peripheral blood and/or CSF from 26 patients suffering from relapsing–remitting MS (17 females and 9 males, aged 19–43 years), who were admitted to the neurological clinic of the University of Rome Tor Vergata for a relapse of the disease. These patients were suffering from a new symptom or a recrudescence of an old one, since at least two days. At the moment of blood and/or CSF withdrawal, patients were free of corticosteroids and MS-specific immunoactive therapies. The diagnosis of relapsing–remitting MS was established by clinical, laboratory and MRI parameters, and matched published criteria (McDonald et al., 2001). In all instances, patients underwent detection of oligoclonal banding in the CSF, and brain and/or spinal cord MRI. Expanded Disability Status Scale (EDSS) scores were between 0.5 and 3.
As controls, we used peripheral blood from 25 age- and gender-matched healthy volunteers (15 females and 10 males, aged 22–41 years), and CSF from 11 subjects (7 females and 4 males, aged 22–47 years) without inflammatory or degenerative diseases of the central or peripheral nervous system. These subjects underwent lumbar puncture because of clinical suspect of acute peripheral neuropathy, meningitis or subarachnoidal haemorrhage. All the subjects gave their written informed consent to the study.
MRI acquisition and analysis
Three Tesla MRI examination during the disease relapse consisted of dual-echo proton density, FLAIR, T2-weighted spin-echo images and pre-contrast and post-contrast T1-weighted spin-echo images. All images were acquired in the axial orientation with 3-mm thick contiguous slices. T2 lesion volume was determined by manual tracing and the number of gadolinium (0.2 ml/kg e.v.)-enhancing lesions was counted by a neuroradiologist who was unaware of the patient's clinical details.
Correlation with endocannabinoid levels was estimated by regression analysis in Prism 3.0 (Graphpad Software Inc., San Diego, CA) and was inspected visually using scatter plots to determine linearity.
EAE mice
As described (Pluchino et al., 2003), chronic, relapsing EAE was induced in C57BL/6 mice by subcutaneous immunization with 300 µl of 200 µg MOG(35–55) (Multiple Peptide System) in incomplete Freund's adjuvant containing 8 mg/ml Mycobacterium tuberculosis (strain H37Ra; Difco). Pertussis toxin (Sigma) (500 ng) was injected on the day of the immunization and again 2 days later. Body weight and clinical score (0 = healthy; 1 = limp tail; 2 = ataxia and/or paresis of hindlimbs; 3 = paralysis of hindlimbs and/or paresis of forelimbs; 4 = tetraparalysis; 5 = moribund or death) were recorded daily.
Mice receiving or not vehicle instead of MOG were employed as controls. Data from these two control groups gave comparable results, so that they were pooled together. All efforts were made to minimize animal suffering and to reduce the number of mice used, in accordance with the European Communities Council Directive of November 24, 1986 (86/609/EEC).
Analysis of the endocannabinoid system
All human and mouse specimens were sent blind to the biochemistry laboratory. For the evaluation of endogenous levels of AEA or 2-AG, lipids were extracted from human CSF or lymphocytes, or from mouse striatal slices, and the organic phase was dried under nitrogen (Maccarrone et al., 2005). Dry pellet was resuspended in 20 µl of methanol, and was processed and analysed by high performance liquid chromatography with fluorimetric detection (Wang et al., 2001), as reported (Maccarrone et al., 2005). In the case of human lymphocytes, blood from 9 control and 9 MS donors (25 ml each) was pooled in three aliquots (3 donors/aliquot), that were used to extract and quantify AEA.
The hydrolysis of [3H]AEA by fatty acid amide hydrolase (E.C. 3.5.1.4; FAAH) was assayed in cell or brain homogenates (10 µg/test), incubated at pH 9.0 with 5 µM [3H]AEA. The release of [3H]arachidonic acid from [3H]AEA was evaluated through reversed phase-high performance liquid chromatography (RP-HPLC), as already reported (Maccarrone et al., 2003). FAAH activity was expressed as pmol [3H]arachidonic acid released per min per mg protein. FAAH expression was determined in lymphocyte extracts by enzyme-linked immunosorbent assay (ELISA), using anti-FAAH polyclonal antibodies (1:250) and GAR-AP (diluted 1:2000), as reported (Maccarrone et al., 2003). Wells were coated with cell extracts (20 µg/well), and the absorbance values at 405 nm of unknown samples were within the linearity range of calibration curves drawn with different amounts of extracts (in the range 0–40 µg/well).
The synthesis of AEA through the activity of N-acyl-phosphatidylethanolamines (NAPE)-hydrolysing phospholipase D (E.C. 3.1.4.4; NAPE-PLD) was assayed in cell or brain homogenates (100 µg/test), using 100 µM [3H]NArPE as reported (Fezza et al., 2005). NAPE-PLD activity was expressed as pmol [3H]AEA released per min per mg protein.
For cannabinoid receptor studies, membrane fractions were prepared from human lymphocytes or mouse brain areas as reported (Maccarrone et al., 2005), and were stored at −80°C for no longer than 1 week. Membrane fractions were used in rapid filtration assays with the synthetic cannabinoid [3H]CP55.940 (400 pM), as described (Gubellini et al., 2002; Maccarrone et al., 2005). The concentration of 400 pM [3H]CP55.940 was chosen on the basis of previous data obtained in similar experimental conditions to avoid saturating effects (Gubellini et al., 2002). In all binding experiments, non-specific binding was determined in the presence of 1 µM ‘cold’ agonist (Maccarrone et al., 2005).
Chemicals used in all biochemical assays were of the purest analytical grade. Anandamide (N-arachidonoylethanolamine, AEA), and 5-(1,1′-dimethyheptyl)-2-[1R,5R-hydroxy-2R-(3-hydroxypropyl)cyclohexyl]-phenol (CP55.940) were purchased from Sigma Chemical Co. (St Louis, MO, USA). N-arachidonoyl-phosphatidyl-ethanolamine (NArPE) was synthesized from arachidonic acid and phosphatidylethanolamine, as reported (Fezza et al., 2005). [3H]AEA (205 Ci/mmol), and [3H]CP55.940 (126 Ci/mmol) were from Perkin–Elmer Life Sciences, Inc. (Boston, MA, USA). [3H]NArPE (200 Ci/mmol) was from ARC (St Louis, MO, USA). Anti-FAAH polyclonal antibodies were prepared by Primm S.r.l. (Milan, Italy) as reported (Maccarrone et al., 2003). Goat anti-rabbit antibodies conjugated to alkaline phosphatase (GAR-AP) were from Bio-Rad (Hercules, CA, USA). N-piperidino-5-(4-chlorophenyl)-1-(2,4-dichloro-phenyl)-4-methyl-3-pyrazole carboxamide (SR141716A) and N-[(1S)-endo-1,3,3-trimethy-1-bicyclo [2.2.1]-heptan-2-yl]5-(4-choro-3-methyl-phenyl)-1-(4-methyl-benzyl)-pyrazole-3-carboxamide (SR144528) were a kind gift of Sanofi-Aventis Recherche (Montpellier, France). Biochemical data reported in this paper are the means ± SD of at least three independent experiments, each performed in duplicate. Statistical analysis was performed by the non-parametric Mann–Whitney U test, elaborating experimental data by means of the InStat 3 program (GraphPAD Software for Science, San Diego, CA).
Electrophysiology on EAE mice
Corticostriatal coronal slices (200 μm) were prepared from tissue blocks of the mouse brain with the use of a vibratome (Centonze et al., 2005). A single slice was then transferred to a recording chamber and submerged in a continuously flowing artificial cerebrospinal fluid (ACSF) (33°C, 2–3 ml/min) gassed with 95% O2- 5% CO2. The composition of the control solution was (in mM): 126 NaCl, 2.5 KCl, 1.2 MgCl2, 1.2 NaH2PO4, 2.4 CaCl2, 11 glucose, 25 NaHCO3.
In both control and EAE brains, the striatum could be readily identified under low power magnification, whereas individual neurons were visualized in situ using a differential interference contrast (Nomarski) optical system. This employed an Olympus BX50WI (Japan) non-inverted microscope with ×40 water immersion objective combined with an infra-red filter, a monochrome CCD camera (COHU 4912), and a PC compatible system for analysis of images and contrast enhancement (WinVision 2000, Delta Sistemi, Italy). Recording pipettes were advanced towards individual striatal cells in the slice under positive pressure and, on contact, tight GΩ seals were made by applying negative pressure. The membrane patch was then ruptured by suction and membrane current and potential monitored using an Axopatch 1D patch clamp amplifier (Axon Instruments, Foster City, CA, USA). Whole-cell access resistances measured in voltage clamp were in the range of 5–20 MΩ.
Whole-cell patch clamp recordings were made with borosilicate glass pipettes (1.8 mm o.d.; 3–5 MΩ), in voltage-clamp mode, at the holding potential (HP) of −80 mV. To study evoked glutamate-mediated excitatory post-synaptic currents (EPSCs), the recording pipettes were filled with internal solution of the following composition (mM): K+-gluconate (125), NaCl (10), CaCl2, (1.0), MgCl2 (2.0), 1,2-bis (2-aminophenoxy) ethane-N,N,N,N-tetraacetic acid (BAPTA; 0.5), N-(2-hydroxyethyl)-piperazine-N-s-ethanesulfonic acid (HEPES; 19), guanosine triphosphate (GTP; 0.3), Mg-adenosine triphosphate (Mg-ATP; 1.0), adjusted to pH 7.3 with KOH. Bicuculline (10 µM) was added to the perfusing solution to block GABAA-mediated transmission. Conversely, to detect evoked GABAA-mediated inhibitory post-synaptic currents (IPSCs), intraelectrode solution had the following composition (mM): CsCl (110), K+-gluconate (30), ethylene glycol-bis (ß-aminoethyl ether)-N,N,N′,N′-tetra-acetic acid (EGTA; 1.1), HEPES (10), CaCl2 (0.1), Mg-ATP (4), Na-GTP (0.3). MK-801 (30 µM) and CNQX (10 µM) were added to the external solution to block, respectively, NMDA and non-NMDA glutamate receptors. For synaptic stimulation, bipolar electrodes were placed on corticostriatal fibres to study EPSCs, and within the striatum to elicit IPSCs. EPSCs and IPSCs were induced at the frequency of 0.1 Hz, while pulse interval was 50 ms (for EPSCs) or 70 ms (for IPSCs) for the experiments on PPR. The electrophysiological traces were stored on P-CLAMP 9 (Axon Instruments) and analysed off-line on a personal computer.
Three to seven cells per animal were recorded. For each type of experiment at least four distinct animals were employed. Throughout the text ‘n’ refers to the number of cells, unless otherwise specified. For data presented as the mean ± SEM, statistical analysis was performed using a paired or unpaired Student's t-test or Wilcoxon's test. The significance level was established at P < 0.05.
Drugs were applied by dissolving them to the desired final concentration in the bathing ACSF. Drugs were: CNQX (10 μM), HU210 (0.3, 1 μM), MK-801 (30 μM), (from Tocris Cookson, Bristol, UK); bicuculline (10 μM) (from Sigma-RBI, St Louis, USA); SR141716A (1 μM) (from Sanofi-Aventis Recherche, Montpellier, France).
Results
To explore the ECS system in MS, we first evaluated the levels of the two endocannabinoids AEA and 2-AG in the CSF of our patients. We found a 6-fold increase of AEA in these subjects (control: 2.4 ± 1.3 pmol/ml; MS: 20.3 ± 15.7 pmol/ml) (n = 11 for both groups, P < 0.001) while 2-AG concentration was normal (n = 5, P > 0.05). Notably, AEA levels were particularly high in those subjects showing contrast-enhancement lesions at the MRI following standard dose (0.2 ml/kg e.v.) of gadolinium (n = 11, R2 = 0.87, P < 0.0001). In contrast, lack of correlation with T2 lesion volume was noted (n = 11, R2 = 0.01, P = 0.74) (Fig. 1).
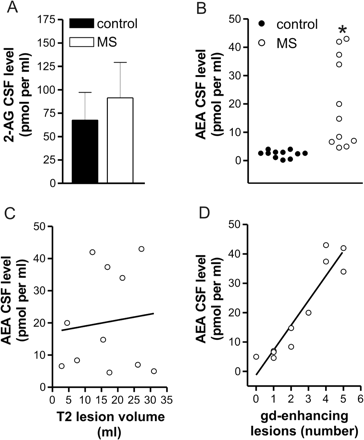
Endocannabinoid levels in the CSF of control and MS subjects. (A) Endogenous levels of 2-AG were unchanged in the CSF of MS patients. (B) This scatter plot shows that endogenous levels of AEA were increased in the CSF of MS patients. (C) The correlation plot shows that AEA levels in MS patients were not related with lesion volume calculated in MRI T2 scans; R2 = 0.01, P = 0.74. (D) The correlation plot shows that AEA levels in MS patients were strongly correlated with the number of active lesions; R2 = 0.87, P < 0.0001. *P < 0.001.
A significant increase of AEA content was also found in peripheral lymphocytes of MS patients (n = 3, P < 0.001). N-acylphosphatidylethanolamine (NAPE)-specific phospholipase D (NAPE-PLD) is considered a major player in the synthesis of AEA (Okamoto et al., 2004), although NAPE-PLD knockout mice do not have altered levels of this endocannabinoid (Leung et al., 2006), possibly due to additional biosynthetic pathways that have been recently identified and await further characterization (Liu et al., 2006). AEA is conversely degradated by the fatty acid amide hydrolase (FAAH) (Cravatt et al., 2001; Maccarrone, 2006). Thus, to see whether the observed increase of AEA was mediated by increased synthesis and/or reduced degradation of this endocannabinoid, we measured the activity of NAPE-PLD and that of FAAH in lymphocytes from the peripheral blood of MS patients. We found that NAPE-PLD activity was increased in MS patients (n = 6, P < 0.0001), while both FAAH activity (n = 17, P < 0.0001) and FAAH protein content (n = 5, P < 0.001) were reduced compared to healthy controls. In peripheral lymphocytes, the binding of CB receptors was conversely normal (n = 16, P > 0.05) (Fig. 2). Incidentally, it should be stressed that AEA levels in peripheral lymphocytes are in the same picomole range as AEA content in human blood (Monteleone et al., 2005), and that FAAH activity and CB receptor binding in healthy subjects corroborate previous data on a large collection of control human lymphocytes (Maccarrone et al., 2001), as does the level of AEA in healthy CSF (Pisani et al., 2005). Furthermore, mammalian immune cells express both CB1 and CB2 receptors (Klein, 2005), and CP55.940 binds both of them (Pertwee, 2002). In human lymphocytes, we found that ∼40% of bound [3H]CP55.940 (400 pM) was displaced by 1 μM SR141716A, a CB1 antagonist (Pertwee, 2002), whereas 1 μM SR144528, an antagonist of CB2 receptors (Pertwee, 2002), displaced it by ∼60% (data not shown). Therefore, it can be concluded that human peripheral lymphocytes express both receptor subtypes to a comparable extent.
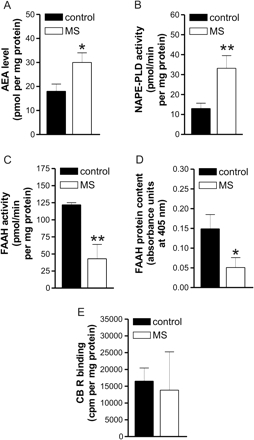
Endocannabinoid metabolism in peripheral lymphocytes of control and MS subjects. (A) AEA levels were increased in peripheral lymphocytes of MS patients. (B) The activity of NAPE-PLD, key enzyme in the AEA synthesis, was increased in MS patients. The activity (C) and protein expression (D) of the AEA degrading enzyme FAAH were reduced in these patients. (E) The binding of CB receptors was conversely unaltered in peripheral lymphocytes of MS patients. *P < 0.001; **P < 0.0001.
Higher concentrations of AEA (n = 3 mice, P < 0.001), coupled with increased activity of NAPE-PLD (n = 4 mice, P < 0.0001) and reduced activity of FAAH (n = 5 mice, P < 0.001), were also observed in the striatum of EAE mice killed 20 to 25 days after EAE induction, at the peak of their neurological deficits (mean clinical score: 2.8). The activity of NAPE-PLD and that of FAAH were conversely normal in the frontal cortex of these mice (Fig. 3). The observed increase in AEA levels extends previous findings on chronic relapsing experimental allergic encephalomyelitis (CREAE) mice, another animal model of MS (Baker et al., 2001). Instead, decreased endocannabinoid levels were reported in the brain of Lewis rats with EAE (Cabranes et al., 2005), suggestive that the animal model of MS (mouse versus rat) might lead to heterogeneity of ECS alterations observed in previous studies. In this context, it should be stressed that the ECS is well regulated and sensitive, and there is the possibility that a number of factors like soluble mediators of inflammation can influence ECS directly or indirectly. Therefore, it is imperative that all experimental conditions are strictly controlled and kept identical, when handling control and EAE mice, in order to minimize possible pitfalls due to differences in the phase of the disorder, or in its association with inflammation. Incidentally, the picomole amounts of AEA found here corroborate previous data in rat striatum (Di Marzo et al., 2000) and mouse brain (Baker et al., 2001).
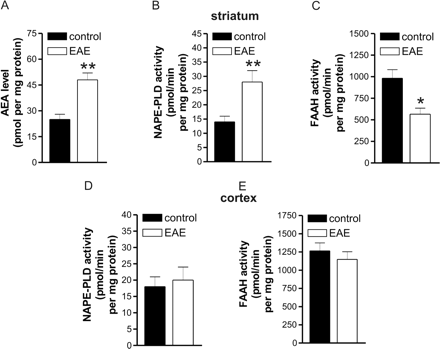
Endocannabinoid metabolism in striatal and cortical slices of control and EAE mice. (A) Endogenous levels of AEA were increased in the striatum of EAE mice. NAPE-PLD activity was increased (B) and FAAH activity was reduced in the striatum of these mice (C). The activity of NAPE-PLD (D) and that of FAAH (E) were conversely normal in the frontal cortex of EAE mice. *P < 0.001, **P < 0.0001.
In EAE mice, we also performed electrophysiological recordings from single neurons to explore the functionality of neuronal ECS. We selected the nucleus striatum for our neurophysiological investigations, since this subcortical area contains among the highest contents of cannabinoid CB1 receptors in the brain (Piomelli, 2003), and it is particularly vulnerable to the neurodegenerative process associated to MS (Bakshi et al., 2002; Bermel et al., 2003).
In the striatum, stimulation of CB1 receptors modulates both excitatory (Gerdeman and Lovinger, 2001; Huang et al., 2001; Gubellini et al., 2002; Centonze et al., 2005) and inhibitory synaptic transmission through a presynaptic action (Szabo et al., 1998; Centonze et al., 2004, 2005). To record glutamate-mediated EPSCs, we performed whole-cell recordings with potassium-based pipettes. At −80 mV HP and in the presence of bicuculline, EPSCs could be detected in striatal neurons following the activation of corticostriatal fibres. Application of the CB1 receptor agonist HU210 dose-dependently reduced EPSCs in control slices (10 min, n = 10 and P < 0.05 compared to pre-drug values for both 0.3 and 1 μM HU210). In striatal neurons from EAE mice, the effects of HU210 were still present and remarkably similar to those recorded in the respective controls (10 min, n = 10 and P < 0.05 for both concentrations). In both experimental groups, the depressant action of HU210 was associated with increased PPR, as expected for a presynaptic action of this compound. Furthermore, pre-incubating the slices with the selective antagonist of CB1 receptors SR141716A failed to change the amplitude of EPSCs, but fully prevented the HU210 effects in both control and EAE mice (for both groups: n = 4 and P > 0.05 for SR141716A and 1 μM HU210) (Fig. 4).
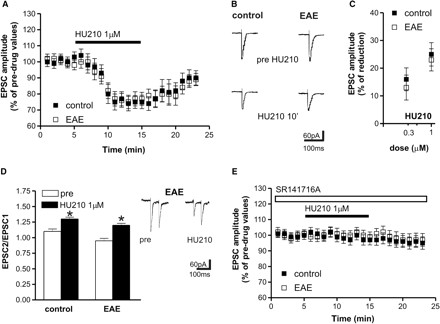
Effects of CB1 receptor stimulation on evoked glutamate-mediated EPSCs in corticostriatal slices of control and EAE mice. (A) Application of the CB1 receptor agonist HU210 reduced EPSC amplitude in control and EAE striatal neurons. (B) Examples of voltage-clamp recordings showing that evoked EPSCs are reversibly reduced by 1 µM HU210. (C) The depressant effect of HU210 was dose-dependent and similar in control and EAE mice. (D) HU210 application enhanced PPR in control and EAE slices. On the right there are samples of PPR recordings before and during the application of HU210 in EAE mice. (E) The depressant action of HU210 was abolished pre-incubating the slices with the selective antagonist of CB1 receptors SR141716A. *P < 0.05.
We also investigated the sensitivity of GABA synapses to HU210. Following intrastriatal stimulation, GABA-mediated IPSCs were recorded, in the presence of MK-801 plus CNQX, in both control and EAE mice. These currents were fully blocked by the GABAA receptor antagonist bicuculline (n = 5 for both groups) and were detected as inward deflections from the baseline. Application of the cannabinoid CB1 receptor agonist HU210 (10 min, n = 8 for both groups and 0.3 and 1 μM HU210) significantly (P < 0.05) reduced IPSCs in control slices, an effect again associated with increased PPR and prevented by SR141716A, which caused, per se, no effect of basal EPSC amplitude in both groups (n = 5 and P > 0.05 for SR141716A and 1 μM HU210). In striatal neurons from EAE mice, HU210 was ineffective (10 min application, n = 10 and P > 0.05 for both concentrations compared to pre-drug values), indicating the loss of CB1-mediated control of GABA transmission in these mice (Fig. 5).
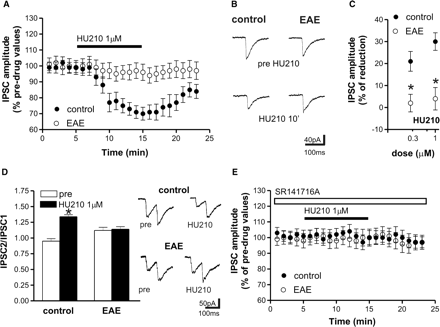
Effects of CB1 receptor stimulation on evoked GABA-mediated IPSCs in corticostriatal slices of control and EAE mice. (A) Application of the CB1 receptor agonist HU210 reduced IPSC amplitude in control but not in EAE striatal neurons. (B) Examples of voltage-clamp recordings showing the lack of effect of HU210 on evoked IPSCs recorded from EAE mice. (C) The depressant effect of HU210 was dose-dependent in control mice. In EAE mice, conversely, the depressant action of HU210 on GABA transmission was lost for both concentrations. (D) HU210 application enhanced PPR in control but not in EAE slices. On the right there are samples of PPR recordings before and during the application of HU210 in control (upper traces) and EAE mice (lower traces). (E) The depressant action of HU210 on IPSC recorded in control mice was abolished pre-incubating the slices with the selective antagonist of CB1 receptors SR141716A. *P < 0.05.
To further explore the possible rearrangements of cannabinoid receptor expression and function in EAE, we measured the binding of the CB1 receptor agonist [3H]CP55,940 to striatal and cortical membranes from EAE and control mice. We found that the binding properties of CB1 receptors were markedly decreased in the striatum of EAE mice (n = 4, P < 0.0001), while this parameter was normal in the cortex (Fig. 6).
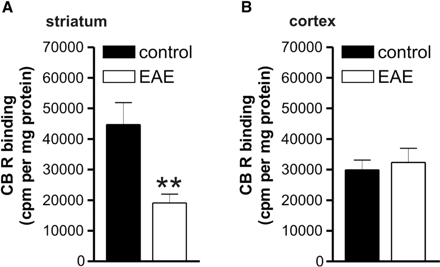
Cannabinoid receptor binding in striatal and frontal cortical slices of control and EAE mice. (A) Cannabinoid CB1 receptor binding was reduced in the striatum of EAE mice. (B) Cannabinoid CB1 receptor binding was conversely normal in the cortex of these animals. **P < 0.0001.
Discussion
The results of the present study show that MS and EAE are associated with significant alterations of the ECS. We found that AEA, but not 2-AG, is increased in the CSF of MS patients, indicating that the two endocannabinoids are differentially engaged during CNS inflammatory diseases. These results are consistent with the emerging concept that AEA and 2-AG, although sharing many pharmacological actions, have differential regulatory mechanisms, and that AEA but not 2-AG is preferentially involved during pathological events (Chevaleyre et al., 2006). We also found increased synthesis, reduced degradation and increased levels of AEA in lymphocytes of MS patients. These data might suggest that an important source of AEA in the CNS of MS subjects are inflammatory cells invading the brain from the periphery. In line with this hypothesis, a previous study reported significantly higher concentrations of AEA in active lesions than in quiescent lesions in MS patients (Eljaschewitsch et al., 2006), and we have observed higher AEA levels in patients with inflammatory activity at the MRI. It should be recalled, however, that because of the special cellular and cytokine microenvironment in the CNS, it is not possible to draw direct conclusions from peripheral lymphocytes to lymphocytes inside the CNS. Furthermore, it should be also noted that only AEA synthesis catalysed by NAPE-PLD was measured in this investigation, through a specific radiochromatographic method that we have recently developed (Fezza et al., 2005) to assay the authentic NAPE-PLD described by Okamoto et al. (2004).
Increased synthesis, reduced degradation and increased levels of AEA have also been detected in the brains of EAE mice in the acute phase of the disease, paralleling the finding in acute MS subjects. In this respect, the normal (Witting et al., 2006) or even low levels of AEA (Cabranes et al., 2005) found in other studies is a likely expression of a different phase of the disease or of a different model. In the study by Witting et al. (2006), in fact, AEA levels have been measured in EAE mice with scarce disease activity (clinical score between 0 and 2), while in the study by Cabranes et al. (2005) EAE has been induced in rats. Furthermore, the consistency of the biochemical data obtained in striatal slices from EAE mice with those seen in patients with MS confirms the validity of our animal model of MS to study critical aspects of the human pathology, and lends support to the conclusion that striatal brain slice preparations do not perturb significantly ECS. In line with this idea, it has been previously observed that the alterations of AEA metabolism in corticostriatal slices from a rat model of Parkinson's disease (Gubellini et al., 2002) were indeed similar to those seen in the CSF of Parkinson's disease patients (Pisani et al., 2005).
In EAE mice, we have also provided direct physiological evidence that neuronal responsiveness to cannabinoid CB1 receptor activation is altered. The selective loss of cannabinoid-mediated control of GABA transmission seen in this model of MS correlates with reduced expression of CB1 receptors, and indicates a fine rearrangement of neuronal ECS, which spares the regulation of glutamate transmission and involves the nucleus striatum, a subcortical area particularly vulnerable to the neurodegenerative process associated to MS (Bakshi et al., 2002; Bermel et al., 2003). This brain region is also a preferential site of action of the ECS in physiological and pathological conditions (Piomelli, 2003). Based on the assumption that the activation of the ECS system in MS and in EAE aims at limiting neuronal damage, it can be hypothesized that the down-regulation of cannabinod receptors inhibiting GABA synapses takes part in this protective action by contrasting neuronal excitotoxic damage.
Excitotoxicity, in fact, plays a crucial role in MS- and in EAE-associated neuronal degeneration, as suggested by several lines of evidence. For example, glutamate clearance is altered in EAE (Hardin-Pouzet et al., 1997; Ohgoh et al., 2002) as well as in MS (Werner et al., 2001; Vallejo-Illarramendi et al., 2006), and glutamate levels are significantly higher in the CSF (Stover et al., 1997; Sarchielli et al., 2003) and in the brains of MS patients (Srinivasan et al., 2005). Finally, glutamate receptor antagonists exert beneficial effects in EAE (Wallstrom et al., 1996; Bolton and Paul, 1997; Pitt et al., 2000; Smith et al., 2000) and in MS (Plaut, 1987), by limiting not only oligodendrocyte but also neuronal damage (Pitt et al., 2000; Smith et al., 2000). In EAE, the neuroprotective effects of cannabinoids are largely dependent on their ability to limit glutamate release and excitotoxic damage through the activation of CB1 receptors (Pryce et al., 2003), implying that the dramatic increase of AEA levels seen in MS patients serves this biological action by stimulating CB1 receptors located on presynaptic glutamatergic nerve terminals. A concomitant stimulation of CB1 receptors regulating GABA release might be detrimental for this anti-glutamatergic action, by reducing in parallel synaptic inhibition. Thus, the selective insensitivity of GABA synapses to cannabinoid-mediated transmitter release seen in acute EAE might be important to prevent this unwanted effect of AEA, and to maximize its anti-glutamatergic action. However, the increase of AEA tone in the brains of EAE mice do not seem to cause maximal neuroprotection, since it has been shown that manipulations that increase further AEA concentrations in EAE result in additional neuroprotective effects (Mestre et al., 2005; Ortega-Gutierrez et al., 2005; Bari et al., 2006; Ligresti et al., 2006). Soluble mediators released during inflammation have been found to dampen AEA elevation in EAE (Witting et al., 2006), possibly explaining why the concentrations of AEA, although elevated, are sub-optimal for this action.
In this study, we have not explored the effects of cannabinoid receptor stimulation on the inflammatory response in MS or EAE. This issue might be relevant to add further information on the biological significance of the observed rearrangement of AEA levels and metabolism in these disorders. However, the existing literature is highly supportive of a beneficial immunomodulatory action mediated by ECS activation during both MS and EAE (Centonze et al., 2007). Accordingly, virtually all immune cells involved in the pathophysiology of MS and EAE express functional cannabinoid receptors (Galiegue et al., 1995; Yiangou et al., 2006), and evidence exists that stimulation of these receptors mediates complex inhibitory actions in these cells, accounting, at least in part, for the protective effects of direct and indirect cannabinoid agonists in MS and animal models of this disease (Lyman et al., 1989; Wirguin et al., 1994; Arevalo-Martin et al., 2003; Croxford and Miller, 2003; Mestre et al., 2005; Ortega-Gutierrez et al., 2005; Ligresti et al., 2006). Accordingly, activation of CB2 receptors inhibits the production of several inflammatory cytokines, inhibits cell proliferation and chemiotaxis, and limits nitric oxide and arachidonic acid release. All these factors are critical determinants for the maintenance of inflammation in MS and EAE (Pertwee, 2002; Walter and Stella, 2004; Ehrhart et al., 2005; Eljaschewitsch et al., 2006; Centonze et al., 2007).
The fact that activated microglia, macrophages and lymphocytes express not only CB2 but also CB1 receptors suggests the possible involvement of both receptor subtypes in the anti-inflammatory effects of endocannabinoids in MS (Facchinetti et al., 2003; Walter and Stella, 2004; present study). In this context, it should be recalled that AEA is a much better agonist of CB1 than CB2 receptors (Sugiura et al., 2000; Pertwee, 2002), therefore it can be suggested that CB1 receptors are more involved than CB2 receptors in inflammatory neurodegenerative disorders like MS, where only the endogenous tone of AEA is modulated. Furthermore, a recent report allowed to extend to non-CB1 and non-CB2 receptors the inhibitory action of cannabinoids on immune cells. It has been demonstrated, in fact, that the synthetic cannabinoid agonist R(+)WIN55,212-2 inhibits the IL-1 signalling pathway in human astrocytes through a pathway which does not involve characterized cannabinoid receptor subtypes (Curran et al., 2005).
In conclusion, together with the previous experimental findings, our results suggest that during immuno-mediated attack of the CNS, the activation of ECS represents a protective mechanism aimed at reducing both neurodegenerative and inflammatory damage through various and partially converging mechanisms that involve neuronal and immune cells.
Acknowledgements
We are grateful to Dr Cinzia Rapino (University of Teramo) for her skillful assistance with biochemical assays. This investigation was supported by Ministero dell’Università e della Ricerca (FIRB 2006) to M.M. and D.C., by Ministero della Salute to M.M. and D.C. (grants 2005 and 2006), by Fondazione TERCAS (Research Programs 2004 and 2005) to M.M. and by Agenzia Spaziale Italiana (DCMC and MoMa projects 2006) to M.M.
References
Abbreviations:
- AEA
anandamide
- 2-AG
2-arachidonoylglycerol
- CB
cannabinoid
- CSF
cerebrospinal fluid
- EAE
experimental autoimmune encephalomyelitis
- ECS
endocannabinoid system
- EPSC
excitatory postsynaptic current
- FAAH
fatty acid amide hydrolase
- HP
holding potential
- IPSC
inhibitory postsynaptic current
- MS
multiple sclerosis
- NAPE-PLD
N-acyl-phosphatidylethanolamines (NAPE)-hydrolysing phospholipase D
- PPR
paired pulse ratio


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}