




Abstract
Asperger’s syndrome (an autistic disorder) is characterized by stereotyped and obsessional behaviours, and pervasive abnormalities in socio‐emotional and communicative behaviour. These symptoms lead to social exclusion and a significant healthcare burden; however, their neurobiological basis is poorly understood. There are few studies on brain anatomy of Asperger’s syndrome, and no focal anatomical abnormality has been reliably reported from brain imaging studies of autism, although there is increasing evidence for differences in limbic circuits. These brain regions are important in sensorimotor gating, and impaired ‘gating’ may partly explain the failure of people with autistic disorders to inhibit repetitive thoughts and actions. Thus, we compared brain anatomy and sensorimotor gating in healthy people with Asperger’s syndrome and controls. We included 21 adults with Asperger’s syndrome and 24 controls. All had normal IQ and were aged 18–49 years. We studied brain anatomy using quantitative MRI, and sensorimotor gating using prepulse inhibition of startle in a subset of 12 individuals with Asperger’s syndrome and 14 controls. We found significant age‐related differences in volume of cerebral hemispheres and caudate nuclei (controls, but not people with Asperger’s syndrome, had age‐related reductions in volume). Also, people with Asperger’s syndrome had significantly less grey matter in fronto‐striatal and cerebellar regions than controls, and widespread differences in white matter. Moreover, sensorimotor gating was significantly impaired in Asperger’s syndrome. People with Asperger’s syndrome most likely have generalized alterations in brain development, but this is associated with significant differences from controls in the anatomy and function of specific brain regions implicated in behaviours characterizing the disorder. We hypothesize that Asperger’s syndrome is associated with abnormalities in fronto‐striatal pathways resulting in defective sensorimotor gating, and consequently characteristic difficulties inhibiting repetitive thoughts, speech and actions.
Received June 5, 2001. Revised November 19, 2001. Second revision February 2, 2002. Accepted February 4, 2002
Introduction
Developmental disorders that lie on the autistic spectrum, including the genetically related subtypes of classical autism and Asperger’s syndrome, are characterized by stereotyped and obsessional behaviours, and pervasive abnormalities in socio‐emotional and communicative behaviour (Wing, 1993, 1996; Gillberg, 1995). Individuals with classical autism have delayed language development, and most have mental retardation (learning disability). Individuals with Asperger’s syndrome have no history of language delay and have normal or superior intellectual abilities, but still show characteristic impairments in reciprocal social interaction (Gillberg, 1998). Thus, in Asperger’s syndrome there is dissociation between cognitive and social skills. However, the neurobiological determinants of the behavioural phenotype of Asperger’s syndrome are poorly understood.
In one of a very few studies of Asperger’s syndrome, Abell et al. (1999) investigated grey matter differences between a group of adults with Asperger’s and matched controls. They noted grey matter anomalies in the cerebellum, and in medial temporal and frontal lobe structures. These findings fit broadly with a growing consensus that limbic system and cerebellar abnormalities may be important determinants of autism. For example, Damasio and Maurer (1978) suggested that developmental abnormalities in a limbic circuit comprising medial prefrontal cortex and temporal lobe, striatum and limbic thalamus underlie autism. This theory has been supported by studies reporting: delayed metabolic maturation (Zilbovicius et al., 1995) and dysfunction of frontal and temporal circuitry (Baron‐Cohen et al., 1999; Critchley et al., 2000; Haznedar et al., 2000; Zilbovicius et al., 2000); decreased correlated metabolic activity between frontal‐parietal and cortical‐subcortical regions (Horwitz et al., 1988); and volume differences in fronto‐temporal regions (Haznedar et al., 1997; Abell et al., 1999; Aylward et al., 1999) and caudate nuclei (Sears et al., 1999). Structural abnormalities in the cerebellum have also been reported in autism, but studies are in disagreement. For example, volume and cross‐sectional area of cerebellar regions have been reported as normal (Piven et al., 1992), increased (Abell et al., 1999) and decreased (Gaffney et al., 1987; Courchesne et al., 1988; Levitt et al., 1999). This failure to replicate most likely results from differences in study methodology and inclusion criteria (e.g. inclusion of both adults and children in the same study, or mixing results from people with and without a learning disability). We therefore restricted the present study to adults with Asperger’s syndrome to clarify neuroanatomical changes in an autistic spectrum disorder uncomplicated by confounds of IQ.
The limbic circuitry, proposed by some as the biological substrate of autism, plays an important role in sensorimotor gating (Bubser and Koch, 1994; Koch and Bubser, 1994; Kodski and Swerdlow, 1995; Swerdlow et al., 1995a; Wan and Swerdlow, 1997; Hazlett et al., 1998; Kumari et al., 1998). We use this mechanism to supress motor responses to irrelevant stimuli, and it is possible that similar processes underlie cognitive gating. A measure of sensorimotor gating is prepulse inhibition of startle (PPI), in which the startle response to a strong stimulus is muted or inhibited when momentarily preceded by a weak stimulus (the prepulse). We hypothesized that people with an autistic spectrum disorder may have defective sensorimotor gating, reflecting their characteristic inability to inhibit or ‘gate’ repetitive thoughts, speech and actions. However, to date, there have been no studies using PPI in adults with Asperger’s syndrome.
Thus there is evidence that people with an autistic spectrum disorder may have abnormalities in brain anatomy, including regions responsible for sensorimotor gating. However, there are relatively few studies of healthy, non‐learning disabled people who lie on this spectrum. We therefore used quantitative MRI in what we believe to be the first comprehensive study of brain anatomy in unmedicated, intellectually able adults with Asperger’s syndrome and healthy controls of comparable age, IQ, gender and handedness. We also carried out a pilot study of sensorimotor gating using PPI. We hypothesized that people with Asperger’s syndrome would have differences in the anatomy and function of limbic circuitry.
Subjects and methods
Subjects
Twenty‐four control adults were recruited locally by advertisement and 21 adults with Asperger’s syndrome were recruited through a clinical research program at the Maudsley Hospital/Institute of Psychiatry (London). All volunteers (see Table 1) gave informed consent (as approved by the Institute of Psychiatry and Bethlem and Maudsley Hospitals Trust research ethics committee), were aged 18–49 years and had an IQ within the normal range (measured using the WAIS‐R, short form; Canavan et al., 1986). None had a history of major psychiatric disorder or medical illness affecting brain function (e.g. psychosis or epilepsy). All had a structured clinical exam and blood tests to exclude biochemical, haematological or chromosomal abnormalities (including fragile X syndrome). Asperger’s syndrome was diagnosed using ICD 10 (International Statistical Classification of Diseases and Health Related Problems—10th revision) criteria (World Health Organization, 1992). In addition, we were able to carry out an autism diagnostic interview (ADI; Lord et al., 1994) in 18 of these individuals who had informants available. All but two reached the ICD 10 diagnostic algorithm cut‐offs for autism in the three domains of impaired reciprocal social interaction (ADIB), communication (ADIC) and repetitive behaviours and stereotyped patterns (ADID). The remaining two people just failed to reach the ADIC algorithm cut‐off.
MRI and analyses
MRI of the brain was carried out on a GE Signa 1.5 Tesla system (General Electric, Milwaukee, Wisc., USA) at the Maudsley Hospital, London. A coronal three‐dimensional spoiled grass (SPGR) data set covering the whole head was acquired (124 slices, 1.5‐mm slice thickness) in addition to a whole brain near axial dual‐echo fast spin echo (FSE) data set aligned with the anterior commissure–posterior commissure (AC‐PC; 3‐mm slice thickness). This enabled two sets of analyses. Manual tracing of brain volumes was performed on the reformatted SPGR dataset using Measure software (Barta et al., 1997), while differences in grey and white matter volumes were determined from the FSE data set using a voxel based Structural Magnetic and Resonance Toolkit (SMaRT) on a SPARC workstation (Sun Microsystems Inc., Surrey, UK; Suckling et al., 1999b; van Amelsvoort et al., 2001; Sigmundsson et al., 2001). All image analysis was carried out blind to subject status.
Manual tracing
As described previously (Murphy et al., 1992, 1993, 1996; van Amelsvoort et al., 2001), we traced the total brain matter volume of cerebral hemispheres and cerebellum, the frontal, parietal, temporal and occipital lobes, and subcortical nuclei (caudate and putamen). We also measured the volume of ventricular CSF. The volume of each region was calculated by multiplying the summed pixel cross‐sectional areas by slice thickness. To control for the relationship of cerebral volume to head size, brain volumes were normalized as a percentage of traced intracranial volume, and analyses were performed on both normalized and raw volumes. Inter‐rater reliabilities were determined for all brain regions traced by the operators and were highly significant (Bartko and Carpenter, 1976).
Voxel‐wise analysis
Voxels representing extracerebral tissue were automatically set to zero (Suckling et al., 1999a) and probability of each intracerebral voxel belonging to grey matter, white matter, cerebrospinal fluid or dura/vasculature tissue classes calculated (Suckling et al., 1999b). Knowing the voxel size (2.2 mm3), the volume of any tissue class could be estimated at each voxel and summed across all intracerebral voxels to yield global tissue class volumes. The images from each FSE data set were co‐registered with a templated image in standard space of Talairach and Tournoux (Press et al., 1992; Brammer et al., 1997) and then applied to each subject’s four‐tissue class probability maps to register them in standard space and allow estimation of between group structural differences (spatial extent statistics) at each intracerebral voxel.
PPI study
Subjects
We studied 12 men from the Asperger’s group and 14 male controls. Two individuals with Asperger’s syndrome and two controls did not generate a startle response and were excluded from further examination. Deficits in PPI are consistently observed in schizophrenia spectrum disorders (Braff et al., 1978; Braff and Geyer, 1990; Braff, 1993; Cadenhead et al., 1993; Perry and Braff, 1994), and others have drawn parallels between schizotypal disorder and autism. Consequently we used the Claridge and Boks Schizotypy questionnaire (Claridge and Boks, 1984) to measure schizotypal traits in all those who participated in this part of our study.
Startle testing
The eyeblink component of the startle response to acoustic stimuli, presented through headphones binaurally, was assessed by recording the electromyogram (EMG) of the orbicularis oculi. Two Ag/AgCl 6‐mm electrodes filled with Dracard electrode paste (SLE, Croydon, UK) were placed under the right eye and a ground electrode was positioned behind the right ear on the mastoid process. A commercially available human startle response monitoring system (Mark II; SR‐Lab, San Diego, Calif., USA) controlled stimulus delivery and EMG recording and scoring. Recording and scoring parameters were identical to those described previously (Kumari et al., 2000). The amplifier gain was kept constant for all subjects, and the EMG was recorded for 250 ms from the onset of the acoustic stimulus (sampling rate 1 ms). The latency to response onset (ms) was defined by a shift of six digital units from the baseline value occurring 18–100 ms after the stimulus. Latency to response peak (ms) was defined as the point of maximal amplitude that occurred within 150 ms from the acoustic stimulus. Responses were rejected if the onset and peak latencies differed by >95 ms or when the baseline values shifted by >90 units.
Test session design
Pulse alone trials comprised a 40 ms pulse stimulus of 116 dB white noise over 70 dB background. The prepulse trials comprised a 20 ms prepulse stimulus of 4 or 16 dB above background, with onset 30 or 120 ms prior to pulse presentation. These four prepulse trials, the pulse‐alone trial and a no‐stimulus trial were presented pseudorandomly in two blocks of 24 trials with a block of six pulse‐alone trials at the beginning and end of the session (60 trials in total). Each block therefore had four of each trial type. The mean inter‐trial interval was 15 s (range 9–23 s). The session began with a 5‐min acclimatization period of 70 dB background noise. Subjects were instructed to look ahead and keep their eyes open.
Statistical analyses
Analysis of MRI data using manual tracing
The analysis of manually traced volumes (Measure) was carried out using SPSS (SPSS 0.8 for Windows, SPSS Inc., Chicago, Illinois, USA). Between‐group differences in total and regional brain volumes were calculated using analysis of covariance (ANCOVA) with group (Asperger’s or control) as the between‐subject variable and age and IQ as covariates. Within‐group age‐related differences in brain volume were explored using Pearson’s correlations. Between group differences in brain ageing were investigated by transforming Pearson’s r into Fisher’s Z‐score to test the significance of the difference between correlations (Pallant, 2001).
Analysis of MRI data using computerized voxel‐wise analysis
The capacity to carry out computerized voxel‐wise analysis (SMaRT) became available later than Measure; therefore, data were available for a subgroup of 17 subjects with Asperger’s syndrome (15 males, two females) and 24 control subjects (22 males, two females). Total grey, white matter and CSF volumes in the Asperger’s and control groups were compared by ANCOVA, with age and IQ as covariates (SPSS 8.0). Significant effects were explored using correlation analysis. However, further analysis of transformed Fisher’s Z‐scores was not possible as there were <20 subjects in the Asperger’s group (Pallant, 2001). Between‐group differences in grey and white matter were localized by fitting an ANCOVA at each intracerebral voxel in standard space as described previously (Bullmore et al., 1999). The level of significance for these spatial extent statistics was set at P < 0.002. At this level of stringency there was less than one false‐positive cluster for each analysis.
PPI
Scores on the Schizotypy questionnaire were compared using a two‐tailed independent sample t‐test. Startle response amplitude was averaged for the six pulse‐alone trials at the beginning and at the end of the session and compared using a two‐tailed independent t‐test. PPI was defined as 100 × (a – b)/a, where, for each of the two blocks, a was the startle amplitude on pulse‐alone trials and b was the startle amplitude on prepulse trials. The impact of Asperger’s on PPI was determined using a 4 (prepulse trial type) × 2 (group) multivariance analysis of variance (MANOVA) with repeated measures over blocks (2). Significant interactions were explored using unpaired Student’s t‐tests (with data collapsed across the session where appropriate). Finally, startle onset latencies were similarly analysed by 5 (pulse, prepulse trial type) × 2 (group) MANOVA with repeated measures over blocks (2). Significant effects were explored using paired Student’s t‐tests (with data collapsed across the session where appropriate).
Results
Brain volumes
Analysis of MRI data using manual tracing
There was no overall effect of group or IQ on any raw or corrected (percentage intracranial volume) manually traced brain volume (Table 2). There was, however, a significant main effect of age on corrected volume of total brain, and both right and left cerebral hemispheres and caudate nuclei (F > 4, P < 0.05 in all cases). In addition, there was a significant group × age interaction in uncorrected volume of total brain, left and right cerebral hemispheres, occipital lobe and caudate nuclei and in corrected volume of whole brain and left hemisphere (F > 4, P < 0.05). The only significant group × IQ interaction noted was that for corrected cerebellar volume [F(1,39) = 5.257, P = 0.03].
The relationship between age and brain volumes was explored using Pearson correlations. In controls, but not in people with Asperger’s syndrome, we found a significant negative relationship between decreasing brain volumes (traced manually) and increasing age (e.g. whole brain volume and age, r = –0.406, P < 0.05; caudate volume and age, r = –0.539, P < 0.05). Moreover, the correlations between age and brain volumes in each group were significantly different (whole brain volume Z = –2.05, P < 0.05; caudate volume Z = –2.66, P < 0.05).
Analysis of MRI data using computerized voxel‐wise analysis
There was no main effect of group or IQ on the total brain volume of grey matter, white matter or CSF derived from computerized voxel‐wise analysis. There was, however, a significant effect of age on CSF volume with both groups having an age‐related increase in CSF volume [F(1,35) = 6.54, P = 0.02]. Additionally, a group × age interaction in grey matter volume approached significance [F(1,35) = 3.59, P = 0.06]. Further exploration revealed a pattern similar to that found by manual tracing; increasing age was significantly correlated with a reduction in grey matter volume in controls (r = –0.406, P < 0.05), but no correlation was found between age and grey matter volume for people with Asperger’s syndrome (Fig. 1).
Spatial extent statistics. The central coordinates and volumes of the three‐dimensional clusters of brain tissues that were significantly different (P = 0.002) are shown in Table 3. Grey matter deficits in subjects with Asperger’s syndrome (Fig. 2) were identified bilaterally in three spatially extensive regions: (i) from the basal ganglia extending to the thalamus and ventral striatum; (ii) within medial frontal lobe and cingulate; and (iii) the cerebellum. The mean volume difference in combined grey matter deficit regions between Asperger’s and control groups was 16% (P < 0.001). No areas of excess grey matter volume were noted in people with Asperger’s syndrome.
White matter deficits in subjects with Asperger’s syndrome were also extensive, but appeared to be concentrated in the left hemisphere (Fig. 3). The deficits extended fronto‐temporally, and from frontal lobe to occipital lobe in left hemisphere apparently within fibre tracts such as the inferior and superior longitudinal fasciculi and occipitofrontal fasciculus. Additional deficits in the pons and left cerebellum were identified. The mean volume difference in combined white matter deficit regions between Asperger’s and control groups was 21% (P < 0.001). In contrast, we identified a bilateral distribution of significant white matter excesses in people with Asperger’s syndrome (Fig. 3); this was concentrated around the basal ganglia, incorporating the external capsule. The mean volume difference in combined areas of white matter excess between Asperger’s and control groups was 42% (P < 0.001).
PPI
There was no significant difference between people with Asperger’s syndrome and controls in mean schizotypy scores [schizotypy score (STA) t(20) = 1.84, schizotypy borderline score (STB) t(20) = 1.72, not significant (ns)], startle amplitude or response onset latency. In both groups, startle reactivity (Table 4) was comparable in first and final blocks (t = –0.16 ns, t = 1.13 ns, respectively). There was no difference in startle onset latencies between groups but response onset latencies decreased in both groups in the 30 ms/4 dB and 30 ms/16 dB conditions compared to the pulse‐alone condition [t(20) = 5.43, P < 0.01; t(20) = 3.99, P < 0.01, respectively].
There was no overall effect of group or block on PPI. However, there was a significant effect of trial type [F(3,18) = 7.51, P < 0.01] and a significant trial × group interaction [F (3,18) = 4.85, P = 0.01]. This was explained by a marked reduction in mean PPI in the Asperger’s group compared with controls in the 120 ms/16 dB prepulse condition [t(20) = –2.68, P = 0.01] (Fig. 4).
Discussion
We believe that this is the first study to quantify MRI differences in both regional brain volumes and grey and white matter volumes in Asperger’s syndrome, and to investigate age related differences in these parameters. Similar to others (Courchesne et al., 1999), we found that megaloencephaly is not a universal feature of autistic spectrum disorders, and we detected no bulk regional brain volume differences between Asperger’s syndrome and controls using manual tracing. Past reports of megaloencephaly in autism (Bailey et al., 1998) may therefore reflect an effect of disease severity (e.g. mental retardation) that is not evident in the Asperger’s sample we studied. We also found that age‐related differences in whole brain and grey matter volume in the controls were not evident in people with Asperger’s syndrome. The reason for this is unknown, but may include neurodevelopmental differences in neurogenesis and programmed cell death.
Our main findings were that people with Asperger’s syndrome had significant reductions in grey matter volume of frontostriatal and cerebellar regions. In addition, people with Asperger’s syndrome had white matter excesses bilaterally around the basal ganglia, whereas they had deficits mainly in left hemisphere. The concentration of abnormalities in frontostriatal circuitry we observed most likely has functional consequences. PPI is thought to depend in part on intact fronto‐striatal pathways (Bubser and Koch, 1994; Koch and Bubser, 1994; Kodski and Swerdlow, 1995; Swerdlow et al., 1995a; Hazlett et al., 1998; Kumari et al., 1998), and we report for the first time a significant impairment in sensorimotor gating in Asperger’s syndrome.
Fronto‐striatal systems in Asperger’s syndrome
Our finding of reduced grey matter in the medial frontal lobe of people with Asperger’s syndrome is in agreement with other neuroanatomical studies of autism (Haznedar et al., 1997; Abell et al., 1999). In contrast, a recent MRI study of basal ganglia size in autism found that volume of the caudate nucleus was increased in subjects with autism and this increase was proportional to an increase in total brain volume (Sears et al., 1999). However, we observed a reduction in grey matter in the basal ganglia in people with Asperger’s syndrome and no increase in caudate or whole brain volume. Important differences between our studies may explain these disparate findings. The Sears et al. (1999) study included children and adults with performance IQs ranging from 52 to 136; the control group had IQs >70. We only studied adults with an IQ >75, and we also identified significant age‐related differences in brain anatomy between people with Asperger’s and controls. Thus our results may differ due to the specific population we studied, in addition to confounds introduced by age and IQ.
Frontal and striatal brain regions are reciprocally connected to each other (Haber et al., 1995) and the thalamus (Ray and Price, 1993). Damasio and Maurer (1978) proposed that dysfunction in a system incorporating the basal ganglia and mesial frontal lobe is responsible for the clinical symptoms of autism, including motor disturbances such as dystonia, bradykinesia and hyperkinesias, and impaired social communication. Their model also predicted temporal lobe abnormalities in autism. Although we did not identify grey matter deficits in the temporal lobe at the level of significance we adopted, we did observe extensive white matter deficits in left temporal lobe. Ours is the first study to report abnormalities in anatomy of this entire neural system in people with an autistic spectrum disorder, and we suggest that our anatomical findings are consistent with Damasio and Maurer’s model.
The fronto‐striatal regions we identified as abnormal are known to have intimate and reciprocal links with cerebellum, and the cerebellum has been implicated in higher order cognitive functions, including executive functions such as planning and shifting attention (Leiner and Leiner, 1989, 1991; Grafman et al., 1992; Appollonio et al., 1993; Courchesne et al., 1994; Middleton and Strick, 1994). Thus, the anomalies we found within the cerebellum, a region anatomically and functionally related to the basal ganglia and frontal cortex, are not surprising. Our study therefore lends tentative support for the hypothesis that abnormalities in the cerebellum may be related to the behavioural phenotype of people with an autistic spectrum disorder, but in our view cerebellar pathology may best be viewed in the context of system‐wide pathology, rather than in isolation.
Sensorimotor gating in Asperger’s syndrome
There is consensus that alterations in fronto‐striatal regions (as found in our study of people with Asperger’s syndrome) underlie impaired sensorimotor gating in a range of neuropsychiatric conditions such as obsessive compulsive disorder (Swerdlow et al., 1993), Huntington’s disease (Swerdlow et al., 1995b), Tourette’s syndrome (Castellanos et al., 1996) and schizophrenia spectrum disorders (Braff, 1993; Cadenhead, 1993). The argument is perhaps most persuasive for Huntington’s disease, as this is associated with significant damage to caudate nucleus (see Klawans and Cohen, 1970). Recent functional imaging data support this position, with activation in prefrontal cortex and caudate nuclei being observed during PPI in healthy individuals (Hazlett et al., 1998; Kumari et al., 1998). We suggest that the reduction in grey matter volume of fronto‐striatal regions we identified in people with Asperger’s may also explain our finding of impaired PPI in this group. Our findings are unlikely to be explained by differences in schizotypal traits, as the control and Asperger’s groups were comparable in this regard.
A previous study of sensorimotor gating in autism (Ornitz et al., 1993) reported no consistent significant differences between diverse groups with autism (comprising adults and children aged 2.8–33 years, IQ 40–145, and some with major medical co‐morbidity) as compared with an intellectually normal healthy control group. In contrast, we explored PPI in a population of intellectually able, healthy, adult men with Asperger’s and found significant impairment of PPI in the 120 ms/16 db condition. Again, we may find differences where others do not due to differences in our study populations.
The occurrence of sensorimotor gating abnormalities in a number of disorders, as noted above, could simply mean that PPI is a task sensitive to fronto‐striatal damage, but differences may not be specific to Asperger’s syndrome. However, sensorimotor gating deficits in autism may reflect similar difficulties with cognitive gating, rendering the individual unable to inhibit or ‘gate’ the repetitive thoughts, speech and actions characteristic of the disorder. The subsequent information ‘overload’ may lead to higher cognitive difficulties, such as executive function and ‘theory of mind’ (ToM) abnormalities, reported by others in autism (Ozonoff et al., 1991; Ozonoff, 1995; Happe and Frith, 1996; Happe et al., 1996). ToM and executive function accounts of autism together explain quite well the socio‐communicative and flexibility problems in this disorder. However, neither account can explain why people with autism are so good at certain tasks. Autism presents a strikingly uneven cognitive profile, with typical peaks on Wechsler block design and digit span. One theoretical account of autism attempts to explain these skills in terms of a bias towards featural versus configural processing. This cognitive style of ‘weak central coherence’ is demonstrated through success of individuals with autism on tasks favouring detail focus, and relative inability on tasks requiring processing of information in context for global form or meaning (Happe, 1996, 1999). Impaired sensorimotor gating permits stimuli indiscriminate access to response output systems without regard for the context of presentation. This is reflected in reduced PPI: the startle response to a given stimulus is not modulated by preceding stimuli. Conceivably, the stimulus which elicits inappropriate startle has been subject to a form of featural processing. We therefore suggest that abnormal gating may contribute to the physiological basis of weak central coherence in autistic spectrum disorders.
White matter deficits and excesses in Asperger’s syndrome
We found widespread white matter anomalies in the brain of people with Asperger’s syndrome; indeed white matter projections to and from abnormal grey matter structures might be expected to be deviant. However, while white matter excesses were distributed bilaterally, deficits appeared to be more prominent in the left hemisphere. This hemisphere normally develops later than the right and, perhaps as a consequence of evolving speech pathways, fronto‐temporal pathways reach maturation later than those linking lower order regions (Paus et al., 1999). Thus neuro‐developmental delay in autism may particularly impact on the left hemisphere and consequently explain some of the developmental language anomalies found in the disorder. For example, we found significant fronto‐temporal white matter deficits in people with Asperger’s syndrome, including the left superior temporal lobe speech area [Brodmann area (BA 22)].
Courchesne et al. (2001) recently investigated developmental changes in grey and white matter volume in autistic boys. They noted that expanded white matter volumes in 2–3 year old children with autism were not found in adolescents with autism. In conjunction with the present findings, this suggests that the autistic brain matures differently, and has a complex and anomalous trajectory affecting both brain development and aging. Courchesne’s group of autistic children had IQs ranging from 36 to 122, while the control group had IQs in the normal range, so it is unclear to what extent the presence of learning disability impacted on their results. Clearly the question of structural brain changes throughout the lifespan of autistic individuals deserves further investigation in well‐matched groups.
Study limitations
Our analysis of structural MRI data involved multiple comparisons thereby introducing a risk of type 1 errors (false‐positive outcomes). However, we feel this is unlikely to fully explain our results. For example, we found significant effects of age on manually traced brain volumes that were consistent with our computerized voxel‐wise analysis. Moreover, in our ANCOVA of the computerized voxel‐wise grey and white matter volumes, the level of significance adopted was chosen specifically to yield less than one false‐positive cluster. Nonetheless, the differential impact of age on brain volumes identified here must be considered only a first step in exploring age‐related differences in brain ageing of people with Asperger’s syndrome. Our study was cross‐sectional and included no children; therefore, we can only report age‐related differences, as opposed to differences in ageing of individuals. Clearly, further longitudinal studies are required. We restricted our study to individuals with Asperger’s syndrome so it is uncertain whether our findings generalize across the entire autism spectrum. Similarly, our study of sensorimotor gating was relatively small and requires replication in larger samples to allow correlations between behavioural and the neurobiological factors. Finally, we did not directly relate our MRI or PPI findings to clinical variables, so we cannot definitively conclude that the anatomical and physiological differences we identified are causally related to the behavioural phenotype of Asperger’s syndrome.
Conclusions
We found that, compared with controls, people with Asperger’s syndrome have age‐related differences in brain anatomy, structural abnormalities in fronto‐striatal systems and the cerebellum, and impaired sensorimotor gating. We suggest that Asperger’s syndrome probably arises from a generalized abnormality in brain development (causing widespread white matter abnormalities). This neurodevelopmental abnormality may, in turn, be modulated by environmental factors such as social isolation. Some regions are more affected than others, and our findings support the hypothesis that a proportion of autistic symptomatology may be explained by frontostriatal disorder. Further studies are required to examine changes in brain anatomy and function across the lifespan, and to explore their relationship to the behavioural phenotype in people with Asperger’s syndrome.
Acknowledgements
The authors wish to thank Mark A. Geyer, Department of Psychiatry, University of California, for his intellectual support of this work in the planning stages and are grateful to two anonymous reviewers for their helpful comments on the manuscript. This study was partly supported by grant from the Wellcome Trust.
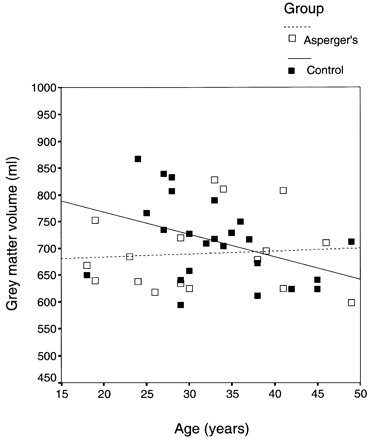
Fig. 1 Age‐related change in grey matter volume in controls and people with Asperger’s syndrome.
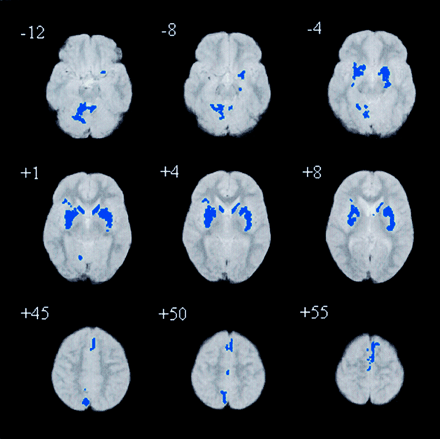
Fig. 2 Relative deficits clusters (blue) in grey matter volume in people with Asperger’s syndrome compared with controls. The maps are orientated with the right side of the brain shown on the left side of each panel. The z coordinate for each axial slice in the standard space of Talairach and Tournoux is given in millimetres.
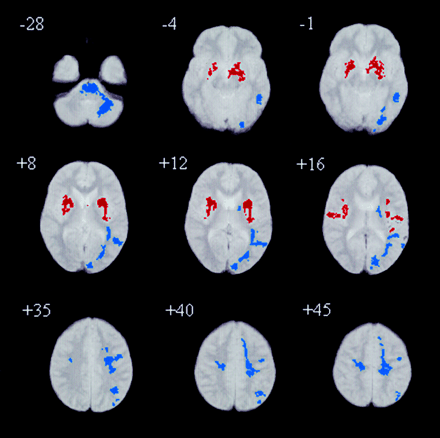
Fig. 3 Relative deficits clusters (blue) and excesses clusters (red) in white matter volume in people with Asperger’s syndrome compared with controls. The maps are orientated with the right side of the brain shown on the left side of each panel. The z coordinate for each axial slice in the standard space of Talairach and Tournoux is given in millimetres.
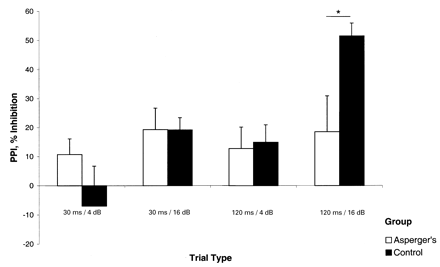
Fig. 4 PPI in Asperger’s syndrome and controls. PPI (± SEM) is shown for each prepulse trial type. PPI is the mean difference between startle response in pulse‐alone trials and prepulse trials, divided by pulse alone trials and multiplied by 100. Compared with controls, people with Asperger’s syndrome have significantly reduced PPI in 120 ms/16 db trials.
Group characteristics
| Group | n | Age (SD) (years) | IQ (SD) | Handedness | Sex | Schizotypy score (range) |
| Asperger’s | 21 | 32 (10) | 96 (15) | 19 right, one left (male) | 19 male, two female | STA 12 (0–21) |
| STB 5 (0–12) | ||||||
| Control | 24 | 33 (7) | 114 (14) | 23 right, one left (male) | 22 male, two female | STA 7 (0–18) |
| STB 3 (0–8) |
| Group | n | Age (SD) (years) | IQ (SD) | Handedness | Sex | Schizotypy score (range) |
| Asperger’s | 21 | 32 (10) | 96 (15) | 19 right, one left (male) | 19 male, two female | STA 12 (0–21) |
| STB 5 (0–12) | ||||||
| Control | 24 | 33 (7) | 114 (14) | 23 right, one left (male) | 22 male, two female | STA 7 (0–18) |
| STB 3 (0–8) |
Age (years) and IQ (full‐scale) values are group means (standard deviation). Schizotypy scores are group means for STA (schizotypy) and STB (borderline) subsections of the Schizotypy questionnaire.
Group characteristics
| Group | n | Age (SD) (years) | IQ (SD) | Handedness | Sex | Schizotypy score (range) |
| Asperger’s | 21 | 32 (10) | 96 (15) | 19 right, one left (male) | 19 male, two female | STA 12 (0–21) |
| STB 5 (0–12) | ||||||
| Control | 24 | 33 (7) | 114 (14) | 23 right, one left (male) | 22 male, two female | STA 7 (0–18) |
| STB 3 (0–8) |
| Group | n | Age (SD) (years) | IQ (SD) | Handedness | Sex | Schizotypy score (range) |
| Asperger’s | 21 | 32 (10) | 96 (15) | 19 right, one left (male) | 19 male, two female | STA 12 (0–21) |
| STB 5 (0–12) | ||||||
| Control | 24 | 33 (7) | 114 (14) | 23 right, one left (male) | 22 male, two female | STA 7 (0–18) |
| STB 3 (0–8) |
Age (years) and IQ (full‐scale) values are group means (standard deviation). Schizotypy scores are group means for STA (schizotypy) and STB (borderline) subsections of the Schizotypy questionnaire.
Whole and regional brain volumes (traced manually) in people with Asperger’s syndrome and healthy controls
| Brain structure | Asperger’s | Control | F | P |
| (n = 21) | (n = 24) | (d.f. = 39) | ||
| Hemispheres | ||||
| % Intracranial volume | ||||
| Total | 74.92 (2.85) | 75.61 (2.71) | 0.03 | 0.87 |
| Left | 37.27 (1.41) | 37.40 (1.36) | 1.18 | 0.28 |
| Right | 37.23 (1.33) | 37.87 (1.25) | 1.89 | 0.18 |
| Raw volume (ml) | ||||
| Total | 1084 (127) | 1105 (118) | 1.49 | 0.23 |
| Left | 539 (61) | 547 (59) | 0.02 | 0.90 |
| Right | 539 (64) | 544 (60) | 0.04 | 0.84 |
| Frontal lobe | ||||
| % Intracranial volume | ||||
| Total | 36.60 (3.03) | 37.68 (1.72) | 0.08 | 0.78 |
| Left | 18.36 (1.92) | 18.59 (0.87) | 0.02 | 0.87 |
| Right | 18.29 (1.56) | 18.87 (1.11) | 0.02 | 0.88 |
| Raw volume (ml) | ||||
| Total | 529 (72) | 550 (61) | 0.08 | 0.78 |
| Left | 266 (46) | 271 (29) | 0.11 | 0.74 |
| Right | 265 (37) | 276 (34) | 0.09 | 0.77 |
| Temporal lobe | ||||
| % Intracranial volume | ||||
| Total | 8.71 (1.02) | 10.74 (5.83) | 2.65 | 0.11 |
| Left | 4.24 (0.56) | 5.16 (2.87) | 1.99 | 0.17 |
| Right | 4.46 (0.53) | 5.56 (2.88) | 3.37 | 0.07 |
| Raw volume (ml) | ||||
| Total | 126 (20) | 156 (77) | 2.48 | 0.12 |
| Left | 62 (11) | 75 (38) | 1.77 | 0.19 |
| Right | 64 (10) | 81 (38) | 3.25 | 0.08 |
| Parietal lobe | ||||
| % Intracranial volume | ||||
| Total | 20.02 (2.27) | 18.16 (2.68) | 0.17 | 0.68 |
| Left | 9.96 (1.08) | 9.06 (1.40) | 0.36 | 0.55 |
| Right | 9.94 (1.16) | 9.08 (1.29) | 0.11 | 0.74 |
| Raw volume (ml) | ||||
| Total | 290 (46) | 266 (49) | 0.08 | 0.78 |
| Left | 144 (21) | 132 (25) | 0.19 | 0.66 |
| Right | 144 (24) | 133 (24) | 0.04 | 0.84 |
| Occipital lobe | ||||
| % Intracranial volume | ||||
| Total | 9.63 (1.97) | 10.26 (2.51) | 0.46 | 0.50 |
| Left | 4.99 (1.12) | 5.30 (1.28) | 1.23 | 0.27 |
| Right | 4.57 (1.01) | 4.92 (1.34) | 0.04 | 0.84 |
| Raw volume (ml) | ||||
| Total | 139 (33) | 150 (38) | 0.26 | 0.62 |
| Left | 72 (17) | 77 (20) | 1.23 | 0.27 |
| Right | 66 (18) | 72 (20) | 0.04 | 0.84 |
| Caudate nucleus | ||||
| % Intracranial volume | ||||
| Total | 0.60 (0.08) | 0.61(0.06) | 1.49 | 0.23 |
| Left | 0.30 (0.04) | 0.31 (0.03) | 1.40 | 0.24 |
| Right | 0.30 (0.04) | 0.30 (0.03) | 1.36 | 0.25 |
| Raw volume (ml) | ||||
| Total | 8.67 (0.98) | 8.76(1.0) | 0.62 | 0.44 |
| Left | 4.31 (0.47) | 4.53(0.67) | 0.67 | 0.42 |
| Right | 4.40 (0.53) | 4.32(0.61) | 0.51 | 0.48 |
| Putamen nucleus | ||||
| % Intracranial volume | ||||
| Total | 0.53 (0.09) | 0.56 (0.07) | 0.03 | 0.86 |
| Left | 0.26 (0.05) | 0.28 (0.04) | 0.001 | 0.97 |
| Right | 0.27 (0.04) | 0.28 (0.04) | 0.15 | 0.70 |
| Raw volume (ml) | ||||
| Total | 7.60 (1.36) | 8.22 (1.25) | 0.005 | 0.94 |
| Left | 3.76 (0.73) | 4.12 (0.64) | 0.02 | 0.90 |
| Right | 3.84 (0.67) | 4.09 (0.65) | 0.08 | 0.79 |
| Lateral ventricles | ||||
| % Intracranial volume | ||||
| Total | 1.60 (1.67) | 1.12 (0.38) | 0.05 | 0.83 |
| Left | 0.88 (1.11) | 0.54 (0.20) | 0.05 | 0.83 |
| Right | 0.82 (0.98) | 0.57 (0.22) | 0.01 | 0.92 |
| Raw volume (ml) | ||||
| Total | 24 (29) | 16 (6) | 0.02 | 0.88 |
| Left | 13.83 (20.56) | 7.94 (3.01) | 0.03 | 0.87 |
| Right | 12.76 (17.64) | 8.30 (3.39) | 0.003 | 0.96 |
| Cerebellum | ||||
| % Intracranial volume | ||||
| Total | 9.06 (1.36) | 9.25 (0.84) | 2.24 | 0.14 |
| Raw volume (ml) | ||||
| Total | 130 (19) | 135 (15) | 1.49 | 0.23 |
| Total cranial volume | 1448 (172) | 1460 (127) | 0.04 | 0.85 |
| Brain structure | Asperger’s | Control | F | P |
| (n = 21) | (n = 24) | (d.f. = 39) | ||
| Hemispheres | ||||
| % Intracranial volume | ||||
| Total | 74.92 (2.85) | 75.61 (2.71) | 0.03 | 0.87 |
| Left | 37.27 (1.41) | 37.40 (1.36) | 1.18 | 0.28 |
| Right | 37.23 (1.33) | 37.87 (1.25) | 1.89 | 0.18 |
| Raw volume (ml) | ||||
| Total | 1084 (127) | 1105 (118) | 1.49 | 0.23 |
| Left | 539 (61) | 547 (59) | 0.02 | 0.90 |
| Right | 539 (64) | 544 (60) | 0.04 | 0.84 |
| Frontal lobe | ||||
| % Intracranial volume | ||||
| Total | 36.60 (3.03) | 37.68 (1.72) | 0.08 | 0.78 |
| Left | 18.36 (1.92) | 18.59 (0.87) | 0.02 | 0.87 |
| Right | 18.29 (1.56) | 18.87 (1.11) | 0.02 | 0.88 |
| Raw volume (ml) | ||||
| Total | 529 (72) | 550 (61) | 0.08 | 0.78 |
| Left | 266 (46) | 271 (29) | 0.11 | 0.74 |
| Right | 265 (37) | 276 (34) | 0.09 | 0.77 |
| Temporal lobe | ||||
| % Intracranial volume | ||||
| Total | 8.71 (1.02) | 10.74 (5.83) | 2.65 | 0.11 |
| Left | 4.24 (0.56) | 5.16 (2.87) | 1.99 | 0.17 |
| Right | 4.46 (0.53) | 5.56 (2.88) | 3.37 | 0.07 |
| Raw volume (ml) | ||||
| Total | 126 (20) | 156 (77) | 2.48 | 0.12 |
| Left | 62 (11) | 75 (38) | 1.77 | 0.19 |
| Right | 64 (10) | 81 (38) | 3.25 | 0.08 |
| Parietal lobe | ||||
| % Intracranial volume | ||||
| Total | 20.02 (2.27) | 18.16 (2.68) | 0.17 | 0.68 |
| Left | 9.96 (1.08) | 9.06 (1.40) | 0.36 | 0.55 |
| Right | 9.94 (1.16) | 9.08 (1.29) | 0.11 | 0.74 |
| Raw volume (ml) | ||||
| Total | 290 (46) | 266 (49) | 0.08 | 0.78 |
| Left | 144 (21) | 132 (25) | 0.19 | 0.66 |
| Right | 144 (24) | 133 (24) | 0.04 | 0.84 |
| Occipital lobe | ||||
| % Intracranial volume | ||||
| Total | 9.63 (1.97) | 10.26 (2.51) | 0.46 | 0.50 |
| Left | 4.99 (1.12) | 5.30 (1.28) | 1.23 | 0.27 |
| Right | 4.57 (1.01) | 4.92 (1.34) | 0.04 | 0.84 |
| Raw volume (ml) | ||||
| Total | 139 (33) | 150 (38) | 0.26 | 0.62 |
| Left | 72 (17) | 77 (20) | 1.23 | 0.27 |
| Right | 66 (18) | 72 (20) | 0.04 | 0.84 |
| Caudate nucleus | ||||
| % Intracranial volume | ||||
| Total | 0.60 (0.08) | 0.61(0.06) | 1.49 | 0.23 |
| Left | 0.30 (0.04) | 0.31 (0.03) | 1.40 | 0.24 |
| Right | 0.30 (0.04) | 0.30 (0.03) | 1.36 | 0.25 |
| Raw volume (ml) | ||||
| Total | 8.67 (0.98) | 8.76(1.0) | 0.62 | 0.44 |
| Left | 4.31 (0.47) | 4.53(0.67) | 0.67 | 0.42 |
| Right | 4.40 (0.53) | 4.32(0.61) | 0.51 | 0.48 |
| Putamen nucleus | ||||
| % Intracranial volume | ||||
| Total | 0.53 (0.09) | 0.56 (0.07) | 0.03 | 0.86 |
| Left | 0.26 (0.05) | 0.28 (0.04) | 0.001 | 0.97 |
| Right | 0.27 (0.04) | 0.28 (0.04) | 0.15 | 0.70 |
| Raw volume (ml) | ||||
| Total | 7.60 (1.36) | 8.22 (1.25) | 0.005 | 0.94 |
| Left | 3.76 (0.73) | 4.12 (0.64) | 0.02 | 0.90 |
| Right | 3.84 (0.67) | 4.09 (0.65) | 0.08 | 0.79 |
| Lateral ventricles | ||||
| % Intracranial volume | ||||
| Total | 1.60 (1.67) | 1.12 (0.38) | 0.05 | 0.83 |
| Left | 0.88 (1.11) | 0.54 (0.20) | 0.05 | 0.83 |
| Right | 0.82 (0.98) | 0.57 (0.22) | 0.01 | 0.92 |
| Raw volume (ml) | ||||
| Total | 24 (29) | 16 (6) | 0.02 | 0.88 |
| Left | 13.83 (20.56) | 7.94 (3.01) | 0.03 | 0.87 |
| Right | 12.76 (17.64) | 8.30 (3.39) | 0.003 | 0.96 |
| Cerebellum | ||||
| % Intracranial volume | ||||
| Total | 9.06 (1.36) | 9.25 (0.84) | 2.24 | 0.14 |
| Raw volume (ml) | ||||
| Total | 130 (19) | 135 (15) | 1.49 | 0.23 |
| Total cranial volume | 1448 (172) | 1460 (127) | 0.04 | 0.85 |
Values are group means (standard deviation). Percentage volumes shown are expressed as a percentage of total cranial volume.
Whole and regional brain volumes (traced manually) in people with Asperger’s syndrome and healthy controls
| Brain structure | Asperger’s | Control | F | P |
| (n = 21) | (n = 24) | (d.f. = 39) | ||
| Hemispheres | ||||
| % Intracranial volume | ||||
| Total | 74.92 (2.85) | 75.61 (2.71) | 0.03 | 0.87 |
| Left | 37.27 (1.41) | 37.40 (1.36) | 1.18 | 0.28 |
| Right | 37.23 (1.33) | 37.87 (1.25) | 1.89 | 0.18 |
| Raw volume (ml) | ||||
| Total | 1084 (127) | 1105 (118) | 1.49 | 0.23 |
| Left | 539 (61) | 547 (59) | 0.02 | 0.90 |
| Right | 539 (64) | 544 (60) | 0.04 | 0.84 |
| Frontal lobe | ||||
| % Intracranial volume | ||||
| Total | 36.60 (3.03) | 37.68 (1.72) | 0.08 | 0.78 |
| Left | 18.36 (1.92) | 18.59 (0.87) | 0.02 | 0.87 |
| Right | 18.29 (1.56) | 18.87 (1.11) | 0.02 | 0.88 |
| Raw volume (ml) | ||||
| Total | 529 (72) | 550 (61) | 0.08 | 0.78 |
| Left | 266 (46) | 271 (29) | 0.11 | 0.74 |
| Right | 265 (37) | 276 (34) | 0.09 | 0.77 |
| Temporal lobe | ||||
| % Intracranial volume | ||||
| Total | 8.71 (1.02) | 10.74 (5.83) | 2.65 | 0.11 |
| Left | 4.24 (0.56) | 5.16 (2.87) | 1.99 | 0.17 |
| Right | 4.46 (0.53) | 5.56 (2.88) | 3.37 | 0.07 |
| Raw volume (ml) | ||||
| Total | 126 (20) | 156 (77) | 2.48 | 0.12 |
| Left | 62 (11) | 75 (38) | 1.77 | 0.19 |
| Right | 64 (10) | 81 (38) | 3.25 | 0.08 |
| Parietal lobe | ||||
| % Intracranial volume | ||||
| Total | 20.02 (2.27) | 18.16 (2.68) | 0.17 | 0.68 |
| Left | 9.96 (1.08) | 9.06 (1.40) | 0.36 | 0.55 |
| Right | 9.94 (1.16) | 9.08 (1.29) | 0.11 | 0.74 |
| Raw volume (ml) | ||||
| Total | 290 (46) | 266 (49) | 0.08 | 0.78 |
| Left | 144 (21) | 132 (25) | 0.19 | 0.66 |
| Right | 144 (24) | 133 (24) | 0.04 | 0.84 |
| Occipital lobe | ||||
| % Intracranial volume | ||||
| Total | 9.63 (1.97) | 10.26 (2.51) | 0.46 | 0.50 |
| Left | 4.99 (1.12) | 5.30 (1.28) | 1.23 | 0.27 |
| Right | 4.57 (1.01) | 4.92 (1.34) | 0.04 | 0.84 |
| Raw volume (ml) | ||||
| Total | 139 (33) | 150 (38) | 0.26 | 0.62 |
| Left | 72 (17) | 77 (20) | 1.23 | 0.27 |
| Right | 66 (18) | 72 (20) | 0.04 | 0.84 |
| Caudate nucleus | ||||
| % Intracranial volume | ||||
| Total | 0.60 (0.08) | 0.61(0.06) | 1.49 | 0.23 |
| Left | 0.30 (0.04) | 0.31 (0.03) | 1.40 | 0.24 |
| Right | 0.30 (0.04) | 0.30 (0.03) | 1.36 | 0.25 |
| Raw volume (ml) | ||||
| Total | 8.67 (0.98) | 8.76(1.0) | 0.62 | 0.44 |
| Left | 4.31 (0.47) | 4.53(0.67) | 0.67 | 0.42 |
| Right | 4.40 (0.53) | 4.32(0.61) | 0.51 | 0.48 |
| Putamen nucleus | ||||
| % Intracranial volume | ||||
| Total | 0.53 (0.09) | 0.56 (0.07) | 0.03 | 0.86 |
| Left | 0.26 (0.05) | 0.28 (0.04) | 0.001 | 0.97 |
| Right | 0.27 (0.04) | 0.28 (0.04) | 0.15 | 0.70 |
| Raw volume (ml) | ||||
| Total | 7.60 (1.36) | 8.22 (1.25) | 0.005 | 0.94 |
| Left | 3.76 (0.73) | 4.12 (0.64) | 0.02 | 0.90 |
| Right | 3.84 (0.67) | 4.09 (0.65) | 0.08 | 0.79 |
| Lateral ventricles | ||||
| % Intracranial volume | ||||
| Total | 1.60 (1.67) | 1.12 (0.38) | 0.05 | 0.83 |
| Left | 0.88 (1.11) | 0.54 (0.20) | 0.05 | 0.83 |
| Right | 0.82 (0.98) | 0.57 (0.22) | 0.01 | 0.92 |
| Raw volume (ml) | ||||
| Total | 24 (29) | 16 (6) | 0.02 | 0.88 |
| Left | 13.83 (20.56) | 7.94 (3.01) | 0.03 | 0.87 |
| Right | 12.76 (17.64) | 8.30 (3.39) | 0.003 | 0.96 |
| Cerebellum | ||||
| % Intracranial volume | ||||
| Total | 9.06 (1.36) | 9.25 (0.84) | 2.24 | 0.14 |
| Raw volume (ml) | ||||
| Total | 130 (19) | 135 (15) | 1.49 | 0.23 |
| Total cranial volume | 1448 (172) | 1460 (127) | 0.04 | 0.85 |
| Brain structure | Asperger’s | Control | F | P |
| (n = 21) | (n = 24) | (d.f. = 39) | ||
| Hemispheres | ||||
| % Intracranial volume | ||||
| Total | 74.92 (2.85) | 75.61 (2.71) | 0.03 | 0.87 |
| Left | 37.27 (1.41) | 37.40 (1.36) | 1.18 | 0.28 |
| Right | 37.23 (1.33) | 37.87 (1.25) | 1.89 | 0.18 |
| Raw volume (ml) | ||||
| Total | 1084 (127) | 1105 (118) | 1.49 | 0.23 |
| Left | 539 (61) | 547 (59) | 0.02 | 0.90 |
| Right | 539 (64) | 544 (60) | 0.04 | 0.84 |
| Frontal lobe | ||||
| % Intracranial volume | ||||
| Total | 36.60 (3.03) | 37.68 (1.72) | 0.08 | 0.78 |
| Left | 18.36 (1.92) | 18.59 (0.87) | 0.02 | 0.87 |
| Right | 18.29 (1.56) | 18.87 (1.11) | 0.02 | 0.88 |
| Raw volume (ml) | ||||
| Total | 529 (72) | 550 (61) | 0.08 | 0.78 |
| Left | 266 (46) | 271 (29) | 0.11 | 0.74 |
| Right | 265 (37) | 276 (34) | 0.09 | 0.77 |
| Temporal lobe | ||||
| % Intracranial volume | ||||
| Total | 8.71 (1.02) | 10.74 (5.83) | 2.65 | 0.11 |
| Left | 4.24 (0.56) | 5.16 (2.87) | 1.99 | 0.17 |
| Right | 4.46 (0.53) | 5.56 (2.88) | 3.37 | 0.07 |
| Raw volume (ml) | ||||
| Total | 126 (20) | 156 (77) | 2.48 | 0.12 |
| Left | 62 (11) | 75 (38) | 1.77 | 0.19 |
| Right | 64 (10) | 81 (38) | 3.25 | 0.08 |
| Parietal lobe | ||||
| % Intracranial volume | ||||
| Total | 20.02 (2.27) | 18.16 (2.68) | 0.17 | 0.68 |
| Left | 9.96 (1.08) | 9.06 (1.40) | 0.36 | 0.55 |
| Right | 9.94 (1.16) | 9.08 (1.29) | 0.11 | 0.74 |
| Raw volume (ml) | ||||
| Total | 290 (46) | 266 (49) | 0.08 | 0.78 |
| Left | 144 (21) | 132 (25) | 0.19 | 0.66 |
| Right | 144 (24) | 133 (24) | 0.04 | 0.84 |
| Occipital lobe | ||||
| % Intracranial volume | ||||
| Total | 9.63 (1.97) | 10.26 (2.51) | 0.46 | 0.50 |
| Left | 4.99 (1.12) | 5.30 (1.28) | 1.23 | 0.27 |
| Right | 4.57 (1.01) | 4.92 (1.34) | 0.04 | 0.84 |
| Raw volume (ml) | ||||
| Total | 139 (33) | 150 (38) | 0.26 | 0.62 |
| Left | 72 (17) | 77 (20) | 1.23 | 0.27 |
| Right | 66 (18) | 72 (20) | 0.04 | 0.84 |
| Caudate nucleus | ||||
| % Intracranial volume | ||||
| Total | 0.60 (0.08) | 0.61(0.06) | 1.49 | 0.23 |
| Left | 0.30 (0.04) | 0.31 (0.03) | 1.40 | 0.24 |
| Right | 0.30 (0.04) | 0.30 (0.03) | 1.36 | 0.25 |
| Raw volume (ml) | ||||
| Total | 8.67 (0.98) | 8.76(1.0) | 0.62 | 0.44 |
| Left | 4.31 (0.47) | 4.53(0.67) | 0.67 | 0.42 |
| Right | 4.40 (0.53) | 4.32(0.61) | 0.51 | 0.48 |
| Putamen nucleus | ||||
| % Intracranial volume | ||||
| Total | 0.53 (0.09) | 0.56 (0.07) | 0.03 | 0.86 |
| Left | 0.26 (0.05) | 0.28 (0.04) | 0.001 | 0.97 |
| Right | 0.27 (0.04) | 0.28 (0.04) | 0.15 | 0.70 |
| Raw volume (ml) | ||||
| Total | 7.60 (1.36) | 8.22 (1.25) | 0.005 | 0.94 |
| Left | 3.76 (0.73) | 4.12 (0.64) | 0.02 | 0.90 |
| Right | 3.84 (0.67) | 4.09 (0.65) | 0.08 | 0.79 |
| Lateral ventricles | ||||
| % Intracranial volume | ||||
| Total | 1.60 (1.67) | 1.12 (0.38) | 0.05 | 0.83 |
| Left | 0.88 (1.11) | 0.54 (0.20) | 0.05 | 0.83 |
| Right | 0.82 (0.98) | 0.57 (0.22) | 0.01 | 0.92 |
| Raw volume (ml) | ||||
| Total | 24 (29) | 16 (6) | 0.02 | 0.88 |
| Left | 13.83 (20.56) | 7.94 (3.01) | 0.03 | 0.87 |
| Right | 12.76 (17.64) | 8.30 (3.39) | 0.003 | 0.96 |
| Cerebellum | ||||
| % Intracranial volume | ||||
| Total | 9.06 (1.36) | 9.25 (0.84) | 2.24 | 0.14 |
| Raw volume (ml) | ||||
| Total | 130 (19) | 135 (15) | 1.49 | 0.23 |
| Total cranial volume | 1448 (172) | 1460 (127) | 0.04 | 0.85 |
Values are group means (standard deviation). Percentage volumes shown are expressed as a percentage of total cranial volume.
Regional differences in grey and white matter volume in people with Asperger’s syndrome and healthy controls (voxel‐based analysis)
| Cluster location | BA | Cluster centre | ||||
| n | x | y | z | Side | ||
| Grey matter deficit | ||||||
| Cerebellum | 217 | 11.1 | –49.8 | –9.2 | Right | |
| Lenticular nucleus (extending to the ventral striatum) | 511 | –23.9 | 1.9 | 4.7 | Left | |
| Lenticular nucleus (extending to the thalamus) | 560 | 28.7 | 1.7 | 7.9 | Right | |
| Cingulate gyrus | 31 | 32 | 1.6 | 50.5 | 28.6 | Right |
| Precuneus | 7 | 36 | 6.7 | –61.3 | 41.9 | Right |
| Precuneus | 7 | 9 | 9.0 | –48.1 | 50.0 | Right |
| Medial frontal gyrus | 6 | 106 | –1.9 | 10.2 | 56.2 | Left |
| Medial frontal gyrus | 6 | 12 | 1.9 | –16.2 | 53.3 | Right |
| Superior frontal gyrus | 8 | 4 | 3.9 | 20.2 | 50.0 | Right |
| White matter deficit | ||||||
| Brain stem | 643 | –11.0 | –36.8 | –23.8 | Left | |
| Frontal, temporal and occipital tracts including: | ||||||
| Inferior longitudinal fasiculus | 775 | –31.0 | –57.7 | 15.6 | Left | |
| Occipitofrontal fasiculus | 522 | –21.3 | –8.7 | 37.8 | Left | |
| Corticospinal tracts | 158 | 12.3 | –20.9 | 53.4 | Right | |
| White matter excess | ||||||
| Basal ganglia/external capsule | 549 | –25.4 | –3.0 | 8.2 | Left | |
| Basal ganglia/external capsule | 424 | 31.8 | 0.3 | 11.3 | Right | |
| Uncinate fasciculus | 4 | –28.3 | 3.9 | –8.0 | Left | |
| Anterior commissure | 3 | –29.2 | –3.4 | –8.0 | Left |
| Cluster location | BA | Cluster centre | ||||
| n | x | y | z | Side | ||
| Grey matter deficit | ||||||
| Cerebellum | 217 | 11.1 | –49.8 | –9.2 | Right | |
| Lenticular nucleus (extending to the ventral striatum) | 511 | –23.9 | 1.9 | 4.7 | Left | |
| Lenticular nucleus (extending to the thalamus) | 560 | 28.7 | 1.7 | 7.9 | Right | |
| Cingulate gyrus | 31 | 32 | 1.6 | 50.5 | 28.6 | Right |
| Precuneus | 7 | 36 | 6.7 | –61.3 | 41.9 | Right |
| Precuneus | 7 | 9 | 9.0 | –48.1 | 50.0 | Right |
| Medial frontal gyrus | 6 | 106 | –1.9 | 10.2 | 56.2 | Left |
| Medial frontal gyrus | 6 | 12 | 1.9 | –16.2 | 53.3 | Right |
| Superior frontal gyrus | 8 | 4 | 3.9 | 20.2 | 50.0 | Right |
| White matter deficit | ||||||
| Brain stem | 643 | –11.0 | –36.8 | –23.8 | Left | |
| Frontal, temporal and occipital tracts including: | ||||||
| Inferior longitudinal fasiculus | 775 | –31.0 | –57.7 | 15.6 | Left | |
| Occipitofrontal fasiculus | 522 | –21.3 | –8.7 | 37.8 | Left | |
| Corticospinal tracts | 158 | 12.3 | –20.9 | 53.4 | Right | |
| White matter excess | ||||||
| Basal ganglia/external capsule | 549 | –25.4 | –3.0 | 8.2 | Left | |
| Basal ganglia/external capsule | 424 | 31.8 | 0.3 | 11.3 | Right | |
| Uncinate fasciculus | 4 | –28.3 | 3.9 | –8.0 | Left | |
| Anterior commissure | 3 | –29.2 | –3.4 | –8.0 | Left |
Location of each three‐dimensional cluster’s centroid in Talairach co‐ordinates [x, y, z (mm)]; n = voxel number in each cluster; BA = Brodmann area. Putative anatomical labels are suggested. Cluster‐wise probability P = 0.002. Two‐dimensional cluster coordinates are not included, see Figs 3 and 4.
Regional differences in grey and white matter volume in people with Asperger’s syndrome and healthy controls (voxel‐based analysis)
| Cluster location | BA | Cluster centre | ||||
| n | x | y | z | Side | ||
| Grey matter deficit | ||||||
| Cerebellum | 217 | 11.1 | –49.8 | –9.2 | Right | |
| Lenticular nucleus (extending to the ventral striatum) | 511 | –23.9 | 1.9 | 4.7 | Left | |
| Lenticular nucleus (extending to the thalamus) | 560 | 28.7 | 1.7 | 7.9 | Right | |
| Cingulate gyrus | 31 | 32 | 1.6 | 50.5 | 28.6 | Right |
| Precuneus | 7 | 36 | 6.7 | –61.3 | 41.9 | Right |
| Precuneus | 7 | 9 | 9.0 | –48.1 | 50.0 | Right |
| Medial frontal gyrus | 6 | 106 | –1.9 | 10.2 | 56.2 | Left |
| Medial frontal gyrus | 6 | 12 | 1.9 | –16.2 | 53.3 | Right |
| Superior frontal gyrus | 8 | 4 | 3.9 | 20.2 | 50.0 | Right |
| White matter deficit | ||||||
| Brain stem | 643 | –11.0 | –36.8 | –23.8 | Left | |
| Frontal, temporal and occipital tracts including: | ||||||
| Inferior longitudinal fasiculus | 775 | –31.0 | –57.7 | 15.6 | Left | |
| Occipitofrontal fasiculus | 522 | –21.3 | –8.7 | 37.8 | Left | |
| Corticospinal tracts | 158 | 12.3 | –20.9 | 53.4 | Right | |
| White matter excess | ||||||
| Basal ganglia/external capsule | 549 | –25.4 | –3.0 | 8.2 | Left | |
| Basal ganglia/external capsule | 424 | 31.8 | 0.3 | 11.3 | Right | |
| Uncinate fasciculus | 4 | –28.3 | 3.9 | –8.0 | Left | |
| Anterior commissure | 3 | –29.2 | –3.4 | –8.0 | Left |
| Cluster location | BA | Cluster centre | ||||
| n | x | y | z | Side | ||
| Grey matter deficit | ||||||
| Cerebellum | 217 | 11.1 | –49.8 | –9.2 | Right | |
| Lenticular nucleus (extending to the ventral striatum) | 511 | –23.9 | 1.9 | 4.7 | Left | |
| Lenticular nucleus (extending to the thalamus) | 560 | 28.7 | 1.7 | 7.9 | Right | |
| Cingulate gyrus | 31 | 32 | 1.6 | 50.5 | 28.6 | Right |
| Precuneus | 7 | 36 | 6.7 | –61.3 | 41.9 | Right |
| Precuneus | 7 | 9 | 9.0 | –48.1 | 50.0 | Right |
| Medial frontal gyrus | 6 | 106 | –1.9 | 10.2 | 56.2 | Left |
| Medial frontal gyrus | 6 | 12 | 1.9 | –16.2 | 53.3 | Right |
| Superior frontal gyrus | 8 | 4 | 3.9 | 20.2 | 50.0 | Right |
| White matter deficit | ||||||
| Brain stem | 643 | –11.0 | –36.8 | –23.8 | Left | |
| Frontal, temporal and occipital tracts including: | ||||||
| Inferior longitudinal fasiculus | 775 | –31.0 | –57.7 | 15.6 | Left | |
| Occipitofrontal fasiculus | 522 | –21.3 | –8.7 | 37.8 | Left | |
| Corticospinal tracts | 158 | 12.3 | –20.9 | 53.4 | Right | |
| White matter excess | ||||||
| Basal ganglia/external capsule | 549 | –25.4 | –3.0 | 8.2 | Left | |
| Basal ganglia/external capsule | 424 | 31.8 | 0.3 | 11.3 | Right | |
| Uncinate fasciculus | 4 | –28.3 | 3.9 | –8.0 | Left | |
| Anterior commissure | 3 | –29.2 | –3.4 | –8.0 | Left |
Location of each three‐dimensional cluster’s centroid in Talairach co‐ordinates [x, y, z (mm)]; n = voxel number in each cluster; BA = Brodmann area. Putative anatomical labels are suggested. Cluster‐wise probability P = 0.002. Two‐dimensional cluster coordinates are not included, see Figs 3 and 4.
Startle reactivity at the beginning and end of PPI test
| Group | Startle amplitude (digital units) | |
| Start | End | |
| Asperger’s | 609 (116) | 491 (118) |
| Control | 632 (95) | 350 (58) |
| Group | Startle amplitude (digital units) | |
| Start | End | |
| Asperger’s | 609 (116) | 491 (118) |
| Control | 632 (95) | 350 (58) |
Values are group means (standard error of the mean).
Startle reactivity at the beginning and end of PPI test
| Group | Startle amplitude (digital units) | |
| Start | End | |
| Asperger’s | 609 (116) | 491 (118) |
| Control | 632 (95) | 350 (58) |
| Group | Startle amplitude (digital units) | |
| Start | End | |
| Asperger’s | 609 (116) | 491 (118) |
| Control | 632 (95) | 350 (58) |
Values are group means (standard error of the mean).
References
Abell F, Krams M, Ashburner J, Passingham R, Friston K, Frackowiak R, et al. The neuroanatomy of autism: a voxel‐based whole brain analysis of structural scans.
Appollonio IM, Grafman J, Schwartz R, Massaquoi S, Hallett M. Memory in patients with cerebellar degeneration.
Aylward EH, Minshew NJ, Goldstein G, Honeycutt NA, Augustine AM, Yates KO, et al. MRI volumes of amygdala and hippocampus in non‐mentally retarded autistic adolescents and adults.
Bailey A, Luthert P, Dean A, Harding B, Janota I, Montgomery M, et al. A clinicopathological study of autism.
Baron‐Cohen S, Ring HA, Wheelwright S, Bullmore ET, Brammer MJ, Simmons A, et al. Social intelligence in the normal and autistic brain: an fMRI study.
Barta PE, Dhingra L, Royall R, Schwartz E. Improving stereological estimates for the volume of structures identified in three‐dimensional arrays of spatial data.
Bartko JJ, Carpenter WT. On the methods and theory of reliability.
Braff DL. Information processing and attention dysfunctions in schizophrenia. [Review].
Braff DL, Geyer MA. Sensorimotor gating and schizophrenia. [Review].
Braff D, Stone C, Callaway E, Geyer M, Glick I, Bali L. Prestimulus effects on human startle reflex in normals and schizophrenics.
Brammer MJ, Bullmore ET, Simmons A, Williams SC, Grasby PM, Howard RJ, et al. Generic brain activation mapping in functional magnetic resonance imaging: a nonparametric approach.
Bubser M, Koch M. Prepulse inhibition of the acoustic startle response of rats is reduced by 6‐hydroxydopamine lesions of the medial prefrontal cortex.
Bullmore ET, Suckling J, Overmeyer S, Rabe‐Hesketh S, Taylor E, Brammer MJ. Global, voxel, and cluster tests, by theory and permutation, for a difference between two groups of structural MR images of the brain.
Cadenhead KS, Geyer MA, Braff DL. Impaired startle prepulse inhibition and habituation in patients with schizotypal personality disorder.
Canavan AG, Dunn G, McMillan TM. Principal components of the WAIS‐R.
Castellanos FX, Fine EJ, Kaysen D, Marsh WL, Rapoport JL, Hallett M. Sensorimotor gating in boys with Tourette’s syndrome and ADHD.
Claridge GS, Boks P. Schizotypy and hemisphere function: I. Theoretical considerations and the measurement of schizotypy.
Courchesne E, Yeung‐Courchesne R, Press GA, Hesselink JR, Jernigan TL. Hypoplasia of cerebellar vermal lobules VI and VII in autism.
Courchesne E, Townsend J, Akshoomoff NA, Saitoh O, Yeung‐Courchesne R, Lincoln AJ, et al. Impairment in shifting attention in autistic and cerebellar patients.
Courchesne E, Muller RA, Saitoh O. Brain weight in autism: normal in the majority of cases, megalencephalic in rare cases.
Courchesne E, Karns CM, Davis HR, Ziccardi R, Carper RA, Tigue ZD, et al. Unusual brain growth patterns in early life in patients with autistic disorder. An MRI study.
Critchley HD, Daly EM, Bullmore ET, Williams SC, van Amelsvoort T, Robertson DM, et al. The functional neuroanatomy of social behaviour: changes in cerebral blood flow when people with autistic disorder process facial expressions.
Gaffney GR, Kuperman S, Tsai LY, Minchin S, Hassaneim KM. Midsagittal magnetic resonance imaging of autism.
Gillberg C. The prevalence of autism and autism spectrum disorders. In: Koot FCVHM, editor. The epidemiology of child and adolescent psychopathology. Oxford: Oxford University Press;
Gillberg C. Asperger syndrome and high‐functioning autism. [Review].
Grafman J, Litvan I, Massaquoi S, Stewart M, Sirigu A, Hallett M. Cognitive planning deficit in patients with cerebellar atrophy.
Haber SN, Kunishio K, Mizobuchi M, Lynd‐Balta E. The orbital and medial prefrontal circuit through the primate basal ganglia.
Happe FG. Studying weak central coherence at low levels: children with autism do not succumb to visual illusions.
Happe F, Ehlers S, Fletcher P, Frith U, Johansson M, Gillberg C, et al. ‘Theory of mind’ in the brain. Evidence from a PET scan study of Asperger syndrome.
Hazlett EA, Buchsbaum MS, Hazneder MM, Singer MB, Germans MK, Schnur DB, et al. Prefrontal cortex glucose metabolism and startle eyeblink modification abnormalities in unmedicated schizophrenia patients.
Haznedar MM, Buchsbaum MS, Metzger M, Solimando A, Spiegel‐Cohen J, Hollander E. Anterior cingulate gyrus volume and glucose metabolism in autistic disorder.
Haznedar MM, Buchsbaum MS, Wei TC, Hof PR, Cartwright C, Bienstock CA, et al. Limbic circuitry in patients with autism spectrum disorders studied with positron emission tomography and magnetic resonance imaging.
Horwitz B, Rumsey JM, Grady CL, Rapoport SI. The cerebral metabolic landscape in autism. Intercorrelations of regional glucose utilization.
Koch M, Bubser M. Deficient sensorimotor gating after 6‐hydroxydopamine lesion of the rat medial prefrontal cortex is reversed by haloperidol.
Kodski MH, Swerdlow NR. Prepulse inhibition in the rat is regulated by ventral and caudodorsal striato‐pallidal circuitry.
Kumari V, Sharma T, Williams SCR, Geyer MA, McAlonan G, Bullmore ET, et al. Functional magnetic resonance imaging of prepulse inhibition of the startle response.
Kumari V, Soni W, Mathew VM, Sharma T. Prepulse inhibition of the startle response in men with schizophrenia. Effects of age of onset of illness, symptoms, and medication.
Leiner HC, Leiner AL, Dow RS. Reappraising the cerebellum: what does the hindbrain contribute to the forebrain. [Review].
Leiner HC, Leiner AL, Dow RS. The human cerebro‐cerebellar system: its computing, cognitive and language skills. [Review].
Levitt JG, Blanton R, Capetillo‐Cunliffe L, Guthrie D, Toga A, McCracken JT. Cerebellar vermis lobules VIII‐X in autism.
Lord C, Rutter M, Le Couteur A. Autism Diagnostic Interveiw – Revised. A revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders.
Middleton FA, Strick PL. Anatomical evidence for cerebellar and basal ganglia involvement in higher cognitive function.
Murphy DG, DeCarli C, Schapiro MB, Rapoport SI, Horwitz B. Age‐related differences in volumes of subcortical nuclei, brain matter, and cerebrospinal fluid in healthy men as measured with magnetic resonance imaging.
Murphy DG, DeCarli C, Daly E, Haxby JV, Allen G, White BJ, et al. X‐chromosome effects on female brain: a magnetic resonance imaging study of Turner’s syndrome.
Murphy DG, DeCarli C, McIntosh AR, Daly E, Mentis MJ, Pietrini P, et al. Sex differences in human brain morphometry and metabolism: an in vivo quantitative magnetic resonance imaging and positron emission tomography study on the effect of ageing.
Ornitz EM, Lane SJ, Sugiyama T, de Traversay J. Startle modulation studies in autism.
Ozonoff S. Executive functions in autism. In: Mesibov ESGB, editor. Learning and cognition in autism. New York: Plenum Press;
Ozonoff S, Pennington BF, Rogers SJ. Executive function deficits in high‐functioning autistic individuals: relationship to theory of mind.
Pallant J. SPSS survival manual. A step by step guide to data analysis using SPSS for Windows (Version 10). Buckingham: Open University Press;
Paus T, Zijdenbos A, Worsley K, Collins DL, Blumenthal J, Giedd JN, et al. Structural maturation of neural pathways in children and adolescents: in vivo study.
Perry W, Braff DL. Information‐processing deficits and thought disorder in schizophrenia.
Piven J, Nehme E, Simon J, Barta P, Pearlson G, Folstein SE. Magnetic resonance imaging in autism: measurement of the cerebellum, pons, and fourth ventricle.
Press WH, Taukolsy SA, Vetterling WT, Flannery BP. Numerical recipes in C: the art of scientific computing. 2nd ed. Cambridge: Cambridge University Press;
Ray JP, Price JL. The organization of projections from the mediodorsal nucleus of the thalamus to orbital and medial pre‐frontal cortex in macaque monkeys.
Sears LL, Vest C, Mohamed S, Bailey J, Ranson BJ, Piven J. An MRI study of the basal ganglia in autism.
Sigmundsson T, Suckling J, Maier M, Williams S, Bullmore ET, Greenwood K, et al. Structural abnormalities in frontal, temporal, and limbic regions and interconnecting white matter tracts in schizophrenic patients with prominent negative symptoms.
Suckling J, Brammer MJ, Lingford‐Hughes A, Bullmore ET. Removal of extracerebral tissues in dual‐echo magnetic resonance images via linear scales‐space features.
Suckling J, Sigmundsson T, Greenwood K, Bullmore ET. A modified fuzzy clustering algorithm for operator independent brain tissue classification of dual echo MR images.
Swerdlow NR, Benbow CH, Zisook S, Geyer MA, Braff DL. A preliminary assessment of sensorimotor gating in patients with obsessive compulsive disorder.
Swerdlow NR, Lipska BK, Weinberger DR, Braff DL, Jaskiw GE, Geyer MA. Increased sensitivity to the sensorimotor gating‐disruptive effects of apomorphine after lesions of medial prefrontal cortex or ventral hippocampus in adult rats.
Swerdlow NR, Paulsen J, Braff DL, Butters N, Geyer MA, Swenson MR. Impaired prepulse inhibition of acoustic and tactile startle response in patients with Huntington’s disease.
vanAmelsvoort T, Daly E, Robertson D, Suckling J, Ng V, Critchley H, et al. Structural brain abnormalities associated with deletion at chromosome 22q11: a quantitative neuroimaging study of adults with velo‐cardio‐facial syndrome.
Wan FJ, Swerdlow NR. The basolateral amygdala regulates sensorimotor gating of the acoustic startle in the rat.
Wing L. The definition and prevalence of autism: a review.
World Health Organization. The ICD 10 classification of mental and behavioural disorders. Geneva: World Health Organization;
Zilbovicius M, Garreau B, Samson Y, Remy P, Barthelemy C, Syrota A, et al. Delayed maturation of the frontal cortex in childhood autism.
Author notes
Departments of 1Psychological Medicine and 2Neuroimaging and 3Social Genetic Developmental and Psychiatry Centre, Institute of Psychiatry, Kings College, 4Functional Imaging Laboratory, Institute of Neurology, 5Department of Psychology, St George’s Hospital Medical School, London, UK and 6Department of Psychiatry, University of Hong Kong, Hong Kong SAR China


{kind=link}
{kind=link}
{kind=link}
{kind=link}