




Abstract
Six subjects with musical hallucinations following acquired deafness are described. The subjects all experienced the condition in the absence of any other features to suggest epilepsy or psychosis. I propose a neuropsychological model for the condition consistent with detailed observation of the subjects' phenomenology. The model is based on spontaneous activity within a cognitive module for the analysis of temporal pattern in segmented sound. Functional imaging was carried out to test the hypothesis that musical hallucinosis is due to activity within such a module, for which the neural substrate is a distributed network distinct from the primary auditory cortex. PET was carried out on the six subjects to identify areas where brain activity increased as a function of the severity of the hallucination. In a group analysis, no effect was demonstrated in the primary auditory cortices. Clusters of correlated activity were demonstrated in the posterior temporal lobes, the right basal ganglia, the cerebellum and the inferior frontal cortices. This network is similar to that previously demonstrated during the normal perception and imagery of patterned–segmented sound, and is consistent with the proposed neuropsychological and neural mechanism.
Introduction
Musical hallucinosis is a disorder of complex sound processing. Subjects perceive complex sound in the form of music in the absence of an acoustic stimulus. As such, the phenomenon might be regarded as an example of mental imagery, defined as `mental acts in which we seem to re-enact the experience of perceiving an object when the object is no longer available' (Halpern and Zatorre, 1999). Mental imagery is, however, usually less vivid than actual perception, and is never attributed to an external process. Musical hallucinosis may be associated with structural brain lesions, epilepsy or psychosis (for reviews, see Berrios, 1990; Keshavan et al., 1992). However, it is most commonly seen in subjects with moderate or severe acquired deafness, and as such it may represent an auditory form of the Charles Bonnet syndrome. This term is more commonly used to describe formed visual percepts in the absence of visual stimulation in subjects with peripheral visual loss (ffytche et al., 1998). This study addresses the basis for musical hallucinosis in six subjects with acquired deafness.
Musical hallucinosis is a disorder of the processing of the high-level pattern in sound. Music comprises discrete sounds that are characterized by fundamental properties such as pitch and onset time. These sounds are built into a high-level pattern at the temporal level of hundreds of milliseconds. Musical hallucinosis requires a mental representation of such a high-level pattern. The mental representation might be based on the same neural mechanism as that active during normal musical perception, although distinct mechanisms for musical perception and imagery are also possible. I use the term `high-level' to refer to structure over and above that in the individual units; in this sense the term might equally be used to refer to tonal or atonal Western music or African or Eastern music.
Previous studies suggest that the processing of discrete sounds depends on a neural substrate different from that used for the processing of high-level patterns formed by these sounds. Several studies suggest that the processing of pitch is related to the function of the human primary auditory cortex in Heschl's gyrus (Zatorre, 1988; Pantev et al., 1989; Griffiths et al., 1998), whilst the onset time of auditory transients is accurately represented in the primary auditory cortex of animals (Phillips and Hall, 1990). Stimulation of the primary auditory cortex at operation is associated with the perception of simple noises or tones (Penfield and Perot, 1963). In contrast, the processing of musical patterns formed by individual sounds is related to a network of cortical areas distinct from the primary auditory cortex. Melody is a pattern of sound pitches, whilst rhythm is a pattern of the onset times and durations of sounds. Studies of melody perception demonstrate a predominantly right-sided network that includes the planum temporale and frontal cortex (Zatorre et al., 1994), whilst the involvement of the cerebellum and basal ganglia has been emphasized in studies of rhythmic processing (Penhune et al., 1998). Stimulation of the superior temporal gyrus outside Heschl's gyrus at operation can be associated with the perception of music (Penfield and Perot, 1963). A study using atonal sequences without conventional rhythm (Griffiths et al., 1999a) suggests the possibility of a general role for superior temporal networks in the analysis of patterns in segmented sound, rather than a specific role in the analysis of music.
This study is a test of the hypothesis that musical hallucinosis in subjects with acquired deafness is related to activity within the neural network for the perception of high-level pattern in sound. The hypothesis makes predictions about the phenomenology of, and the brain substrate for, musical hallucinosis. In terms of phenomenology, the hypothesis predicts that subjects with musical hallucinosis due to acquired deafness will experience normal high-level sound patterns because of activation of the normal central perceptual mechanism. In terms of brain substrate, the hypothesis predicts that musical hallucinosis will be associated with activity in distributed networks, distinct from the primary auditory cortex, including the posterior superior temporal cortex, frontal cortex and cerebellum. I was particularly interested to examine whether the planum temporale is active during musical hallucinosis. Several studies suggest a general role for this area in the perception of patterned–segmented sound (Binder et al., 1996; Mummery et al., 1999).
Patients and methods
Table 1 gives the clinical details for the subjects, all of whom were initially referred to neurologists with the exception of Subject 5, who was initially referred for a psychiatric opinion. All subjects gave informed consent to take part in this study, which was conducted with the approval of the ethical committee of the National Hospital for Neurology and Neurosurgery (London).
ENT symptoms/audiometry
All subjects had moderate or severe bilateral deafness, as assessed by pure-tone audiometry (Fig. 1). All subjects suffered progressive hearing loss. No subject had a family history of deafness. In Subject 2 the symptomatic onset occurred when the subject was working as a tank crew member, whilst in Subject 5 there were two episodes of acute lateralized hearing loss following head injuries. The duration of symptomatic hearing loss varied between 5 and 40 years. Subjects 4 and 5 experienced accompanying tinnitus whilst Subjects 2 and 4 had experienced rotatory vertigo (related to head movement in Subject 2, and in only the early history in Subject 4). No subject experienced a syndrome to suggest Menière's disease (fluctuating deafness, fluctuating tinnitus, fluctuating vertigo or aural fullness).
Case histories
The subjects perceived music in the absence of any musical stimulus. Several of the subjects had initially thought that actual music was playing, but all subjects subsequently attributed the experience to a problem with the brain or the ears (the nature of the percept did not change). Apart from low mood accompanying the experiences, which all of the subjects found distressing, no subject described any accompanying delusion. There was no description of experiential features (Gloor, 1990) or generalized convulsions in any subject. The abnormal auditory experiences were restricted to the musical domain in all of the subjects except Subject 6, who also experienced auditory hallucinations in the form of speech and environmental sounds. All of the subjects described an experience of continuous or near-continuous musical hallucinations and described variation in the severity of their symptoms over the course of the day (which was an inclusion criterion for the study). There was no historical suggestion in any of the subjects of any accompanying difficulty with the perception of music, speech or environmental sound over and above that due to the deafness; I did not feel that any of the subjects had an apperceptive auditory agnosia (Griffiths et al., 1999b). One of the subjects (Subject 1) had noticed a small decrease in symptoms with the consistent use of a hearing aid. None of the other subjects, all of whom had used amplification devices, felt that these had any effect on the severity of the experience.
Subject 1
This subject, with 5 years of symptomatic hearing loss, has been reported previously as a single case (Griffiths et al., 1997). He had a history of <1 year of continuous musical hallucinations in the form of multiple singers singing familiar melodies with indistinguishable lyrics. The onset was abrupt and not related to any other symptoms. The songs included hymns and rugby songs, and recent popular music.
Subject 2
This subject, with approximately 40 years of symptomatic hearing loss, had a history of 3 years of continuous musical hallucinations. The onset was abrupt and was not accompanied by other neurological symptoms. The songs included light operatic pieces, and popular songs by artists including Shirley Bassey and Boyzone.
Subject 3
This subject, with 40 years of symptomatic hearing loss, had a history of 9 years of almost continuous musical hallucinations. The onset was abrupt and not accompanied by other symptoms. The experience would usually take the form of organ or piano music, which might be accompanied by singers. If accompanied by singers, the lyrics would be distinguishable. The songs included hymns, nursery rhymes and old popular songs.
Subject 4
This subject, with 23 years of symptomatic hearing loss, had a history of more than 10 years of almost continuous musical hallucinosis. The onset of the hallucinations was not abrupt. The experience was different from that of the other subjects in that she did not hear instruments or singers but experienced musical hallucinations in which the individual notes had the quality of a buzzy pitch. In psychoacoustic terms her experience would be equivalent to hearing notes with pitch, where the individual pitches were associated with band-pass noise with different passbands. Her experience had evolved from almost continuous tinnitus with similar characteristics to the buzzy pitches experienced as music. She currently chooses not to listen to actual music because it is so distorted at the level she needs to listen.
Subject 5
This subject, with more than 40 years of progressive deafness, had a history of 2 years of continual musical hallucinations in the form of three or four male singers singing familiar songs with accompanying musical instruments. The onset followed shortly after a head injury, and was accompanied by hearing a localized noise behind his head and experiencing a hot feeling that rose up the back of his head on two occasions only. There were no accompanying experiential features or depression of his conscious level. The songs usually dated from before the 1970s, when the patient lost interest in listening to music. He finds the sound of actual music distorted.
Subject 6
This subject, with 15 years of progressive deafness, had a history of less than 3 years of continual musical hallucinations. The onset of the events was coincident with an episode when she developed loss of vision, disorientation, perplexion, slurring of her speech and unsteadiness of body; this is likely to have been a posterior circulation vascular event. She would hear one or more singers and accompanying music in the form of a piano or band. Unlike the five other subjects, she also experienced environmental sound hallucinations and verbal hallucinations. The environmental sound hallucinations included wartime planes and sirens, and the sound of dogs barking and children crying. The verbal hallucinations included hearing indistinguishable sounds similar to the murmuring of a crowd and the sound of her sisters talking to each other. The voices never talked about her or to her. She also experienced palinacousis; she described several prolonged hallucinations in the form of hymns triggered by listening to the television programme `Songs of Praise'.
Structural imaging
All subjects underwent structural imaging using MRI (Subjects 1–5) or CT (Subject 6). No subject demonstrated loss of grey or white matter volume. MRI of Subjects 2 and 4 demonstrated multiple areas of signal change in the deep white matter of the cerebral hemispheres consistent with small vessel disease (Subject 2, Fig. 2). Subject 6 had a large arachnoid cyst in the right occipital lobe (Fig. 2).
Functional imaging
Functional imaging was carried out using PET to measure regional cerebral blood flow as a measure of local cerebral activity. A parametric design was used to demonstrate areas of the brain where activity varied as a function of the reported severity of the hallucinosis (Silbersweig et al., 1995; Griffiths et al., 1997). For each subject, 12 PET scans were performed on a Siemens scanner with 3D acquisition using the intravenous oxygen 15 water bolus technique to estimate regional cerebral blood flow. Twelve acquisitions were performed in two separate blocks of six that were 12 h apart. This was in order to maximize the differences in symptom severity corresponding to the scans. Before the onset of each scan, the subjects were told that a scan was about to be carried out and asked to close their eyes. At the end of each scan they were told that the scan had finished and asked to give a rating, between 1 and 7, of the severity of musical symptoms during the scan. A rating of 1 corresponded to no perception of music and 7 to the most severe experience ever. Subjects also reported the features of the experience after each scan in response to structured questions, as shown in Table 2. There was no output task during the scan.
PET image processing and statistical analysis were carried out using statistical parametric mapping software (SPM99b, http://www.fil.ion.ucl.ac.uk/spm). Scans were realigned and spatially normalized (Friston et al., 1995) to the standard stereotaxic space of Talairach and Tournoux (Talairach and Tournoux, 1988). The data were smoothed with a Gaussian filter (filter width at half maximum, 16 mm). Analysis of covariance was used to correct for differences in global blood flow between the scans and to implement individual and group regression analyses to find areas where blood flow increased with symptom severity. The significance of this regression was assessed with the t statistic at each voxel. These statistics (after transformation to a Z score) constitute a SPM{Z}.
Results
Systematic study of phenomenology
Systematic interrogation of the subjects every 8 min over the course of the PET experiment yielded the features reported in Table 2. All of the subjects showed variation in the severity of the hallucinosis except for Subject 5, in whom the severity was fixed. All subjects reported hearing normal patterns of pitch and rhythm that formed recognizable tunes consistent with their listening experience and interests. Only Subjects 1 and 2, who had the least severe hearing loss (~50 dB), experienced contemporary tunes. In a single scan for one subject, the reported experience was of music without a recognizable tune, although it had similar characteristics to other pieces heard during scanning. All of the subjects, except Subject 4, heard singers during their imaging, with or without distinct lyrics. Four of the subjects heard distinct or indistinct instruments.
PET study to demonstrate areas of activity correlated with hallucinosis
Individual analyses were carried out on each of the six subjects except Subject 5, who failed to show any variation in the severity of the hallucinations over the course of the experiment. In Subjects 1–4, the analyses were carried out on realigned, normalized and smoothed data. In Subject 6 the analysis was carried out on realigned and smoothed data, because of the markedly abnormal brain structure. The individual analyses were carried out primarily to seek activation in the primary auditory cortex in Heschl's gyrus (HG) as a linear function of hallucinosis strength, taking HG as the a priori region of interest. The centre of mass of HG from the probabilistic map of Penhune and colleagues (Penhune et al., 1996) (left coordinates –45, –20, 8; right coordinates 45, –15, 5) was used to define the centre of a spherical volume of interest within HG of diameter 1 cm. Within this volume, two subjects showed significant activation at the P < 0.05 voxel level with correction for the size of the volume of interest: Subject 2 for the left HG (P < 0.001 corrected voxel level) and Subject 3 for the right HG (P < 0.005 corrected voxel level). Analysis was also carried out on the individual data sets to seek activation as a function of hallucinosis strength in the planum temporale (PT) as a region of a priori interest. The centre of mass of the PT from the probabilistic map of Westbury and colleagues (Westbury et al., 1999) (left coordinates –60, –30, 10; right coordinates 65, –30, 10) was used to define the centre of a spherical volume of interest within the PT of diameter 1 cm. Three subjects showed significant activation within this volume at the P < 0.05 voxel level with correction for the size of the volume of interest. Significant activation was demonstrated in the three subjects as follows: Subject 1 (left PT, P < 0.05 corrected); Subject 2 (right PT, P < 0.05 corrected); Subject 3 (right PT, P < 0.005 corrected).
Group analysis was carried out for Subjects 1–4 (Table 3 and Figs 3 and 4). Subject 5 was excluded from the analysis because of lack of variation in the severity of hallucinosis, and Subject 6 was excluded from the analysis because of markedly abnormal brain structure. The group analysis was carried out to demonstrate the typical behaviour of the group. Formally, this represents a fixed-effects analysis. The four subjects' similar experiences during scanning, age, handedness and musicality justify such an analysis. No activation as a function of hallucinosis strength was demonstrated in HG on either side in the group analysis. Using the same volume-of-interest method for HG as in the individual analyses, no activation was shown at the P < 0.05 level with correction for the size of the volume. Using the same method for PT as in the individual analyses, significant activation as a function of hallucinosis strength was shown in the left and right plana temporale (left P < 0.005, right P < 0.05, corrected voxel level). The activation in each planum temporale (Fig. 4) formed part of a cluster extending on to the lateral surface of the posterior superior temporal gyrus, as shown in Fig. 3. In the whole-brain analysis, without taking any prior hypotheses into account (Table 3 and Figs 3 and 4), significant clusters of activation were demonstrated in the right basal ganglia and right frontal operculum, the posterior temporal lobes (especially the right), both lobes of the cerebellum, the left deep sylvian cortex and the left frontal lobe.
Discussion
In this study I observed the detailed phenomenology of musical hallucinosis in six patients with acquired deafness. The subjects all had musical hallucinations that occurred after the onset of deafness in the absence of features to suggest psychosis or epilepsy. Based on the phenomenology, I will argue for a model for the production of musical hallucinations in such patients based on activity within a common mechanism for the perception of pattern in segmented sound. The functional imaging data suggest a neural substrate for this common mechanism.
Model
Figure 5 shows a model for the normal and abnormal perception of patterned–segmented sound. This is a cognitive neuropsychological model (Ellis and Young, 1988) based on modular psychological mechanisms, although I will also argue for the likely neural substrate for its implementation.
During normal listening to patterned–segmented sounds such as music, the auditory input is processed by two perceptual mechanisms, operating in a hierarchical fashion (Fig. 5, top). The perceptual mechanism for individual sounds operates before that for the perception of the pattern formed by these sounds. After the pattern has been perceived, it is encoded into memory. The model is based on a single module being active during both the perception and the imagery of patterned sound. In acquired deafness there is impoverished normal input to this module. I propose that this allows spontaneous activity within the module (Fig. 5, bottom). I also propose that the recognition system for patterned sound can interact strongly and reciprocally with the perception/imagery module in the absence of normal processing activity in the latter. This may produce positive feedback, represented by the bold arrows in both directions in the lower part of Fig. 5, between the perception/imagery module and encoding/recognition modules.
The model predicts first that musical hallucinosis due to acquired deafness will be associated with the perception of normal temporal patterns within segmented sound that are consistent with the subject's previous experience. This is the case in all six subjects observed. In the model, this is the result of the interaction between the perception/imagery and recognition modules, the latter accessing patterns encoded during previous listening experience. All of the subjects experienced music that had been heard before they developed severe deafness, and it is notable that Subjects 1 and 2, who had experienced more contemporary music, have less severe hearing loss than the others. In the subjects with the most marked hearing loss (Subjects 4 and 5), there was a disparity between the normal experience of old tunes during hallucinosis and the distorted perception of actual music; this is entirely consistent with the model. Subject 4 is of particular interest with respect to her normal perception of temporal pattern during hallucinosis. She perceives normal temporal patterns of pitch, although the individual pitches are not associated with normal musical notes. This is consistent with distinct mechanisms for the processing of the features of individual sounds compared with the processing of higher-level patterns formed by these sounds. Essentially, I am arguing in her case that abnormal activity in the module for the perception of individual sounds can have a normal higher-order structure imposed by the perception/imagery module. Some studies of tinnitus (Lockwood et al., 1998) suggest low-level activity in the individual sound module (which, I have argued, is the primary auditory cortex), whereas others suggest more distributed high-level processing (Giraud et al., 1999). This difference might reflect whether the tinnitus has associated high-level temporal or spatial structure associated with it.
Secondly, the model allows for the `triggering' of activity in the perception/imagery module by the impoverished auditory input. For some sounds the low signal-to-noise ratio in such input might lead to misperception and misrecognition of certain incoming sounds as music followed by mental `amplification' due to the positive feedback loop between the perception/imagery and recognition modules. The model also allows for the triggering of activity in the perception/imagery module by any abnormal activity from the module for the perception of individual sounds. Gordon has particularly stressed the possible contribution of early parts of the auditory pathway to musical hallucinations (Gordon, 1994). I would point out here only that the proposed model does not require the ascending auditory system to represent a true musical pattern to produce the phenomenon of musical hallucinosis. Triggered activity would also provide a mechanism for the phenomenon of palinacousis seen in Subject 6.
Two questions arise immediately from the model. First, the model is based on the processing of patterned sound rather than music per se. The question arises of why the subjects do not usually perceive other forms of patterned–segmented sound, such as speech. Speech, like music, contains a high-level structure at the level of hundreds of milliseconds, called prosody or the `melody of language' (Monrad-Krohn, 1947). Speech, unlike music, is also characterized by spectral and temporal complexity within the segmented sounds (in terms of temporal structure, at the level of milliseconds and tens of milliseconds). The production of verbal hallucinosis in this model would therefore require additional abnormal activity in the module for the perception of individual sounds, as well as in that for the perception of higher-order pattern. This is possible within the model, although less likely. Only one of the six subjects in this study also suffered verbal hallucinations in the form of spoken speech, although in 41 of the 72 scans observed the subjects perceived speech with accentuated pattern or prosody in the form of singing. Previous studies in a broader population, including subjects with psychiatric disorders, epilepsy and structural lesions (Berrios, 1990), have also reported verbal hallucinations in subjects with musical hallucinations. As in the present study, associated auditory verbal hallucinations in the form of spoken speech were found only in a minority of subjects. A second question is why all subjects with acquired deafness of the degree shown by these subjects do not experience musical hallucinosis, if this is due to the activity of a normal module cut off from its normal input? This might reflect differences between subjects in a threshold for spontaneous or triggered activity within the perception/imagery module.
Model implementation
In terms of the neural mechanisms that might underlie this model, I argued earlier that the perception of the features of individual sounds involved neural mechanisms at or close to the primary auditory cortex in HG, whilst the perception of higher order patterns formed by such sounds involved more widely distributed mechanisms, including the planum temporale. A prediction of the model is therefore that increasing intensity of hallucinosis with increasing spontaneous activity in the module for pattern perception will be associated with increasing activation in a network distinct from HG, including the planum temporale. The group analysis has demonstrated that the typical behaviour of the group is to show such increasing activation with increasing hallucinosis strength in the planum temporale. The typical behaviour of the group is not to show such activation in HG, although it was shown in one or other HG in two of the individual analyses.
A striking feature of this study is the similarity of the activation produced by musical hallucinosis when there is perception without input and the perception of actual patterned–segmented sound in controls (Fig. 4). This is consistent with the proposed model. However, the perception of actual patterned–segmented sound also leads invariably to activation in the primary auditory cortex, something that has not been demonstrated in this study of hallucinosis. The analysis of hallucinating patients is not consistent with the primary auditory cortex being a sufficient substrate for higher-order pattern perception. The model in Fig. 5 does not make strong predictions about activation in the module for the perception of individual sounds in hallucinosis, but this could occur with descending excitation from the pattern-perception module to the module for the perception of individual sounds. If this were the mechanism, the present data suggest that such feedback activation is not universal.
Activation in a cortical network including the right planum temporale has been shown in previous studies of the basis of musical perception (Zatorre et al., 1994). Moreover, studies of musical imagery have demonstrated a network similar to that for perception (Zatorre et al., 1996; Halpern and Zatorre, 1999), which supports the common mechanism for musical perception and imagery proposed in this model. This network includes predominantly the right auditory association cortex (including the planum temporale) and the right and left frontal cortices.
The model I propose is based on activity within a module for the perception and imagery of pattern within segmented sound rather than music per se. Imaging studies that incorporate a task which demands imagery of simple auditory patterns (Rao et al., 1997; Penhune et al., 1998) have also demonstrated activation of the right planum temporale in the absence of activity in the primary auditory cortex. This is consistent with the activation of the planum temporale in the current study being a mechanism for the perception and imagery of patterned–segmented sound, the primary auditory cortex having an obligatory role in perception only.
The role of central lesions
The role of central lesions in producing musical hallucinations has been the subject of controversy. Although gross structural lesions in the brainstem or either hemisphere can be associated with musical hallucinosis (Tanabe et al., 1986; Paquier et al., 1992; Inzelberg et al., 1993; Murata et al., 1994) such lesions are not seen in most patients with this condition. In the present study, one subject had a large structural anomaly (arachnoid cyst), of uncertain relevance, in addition to a major-territory vascular event. Two other subjects had multiple vascular lesions affecting the white matter, whilst two others had vascular risk factors (in addition to age) without demonstrated lesions on MRI.
Central vascular lesions might be the additional factor that distinguishes subjects with musical hallucinosis from subjects with the same degree of deafness without hallucinosis. Several mechanisms are possible. First, it is possible that diffuse vascular disease might lead to a degree of disconnection between the primary auditory and association cortices. In such a way the input to the perception/imagery module might be more diminished in the group with vascular disease. A second possibility is that a vascular lesion or lesions within the network for the perception and imagery of segmented sound alters the threshold for spontaneous activity within this network. I favour the second mechanism, as the distributed nature of the network would make it more likely to be affected by a stochastic process such as small vessel disease. Moreover, the second mechanism involves a network that includes deep brain structures, which are more likely to be affected by small vessel disease than the superior temporal cortex. For either mechanism, vascular disease would act as a factor to augment the suggested process in the model dependent on peripheral deafness. Such vascular disease would not be an adequate explanation in itself.
Conclusion
I have proposed a model for the production of musical hallucinations in subjects with acquired deafness in the absence of psychosis or epilepsy. This model is based on spontaneous activity in a module usually involved in the normal perception of pattern in segmented sounds. Functional imaging data support the hypothesis that such a module is physically realized in a distributed cortical network distinct from the primary auditory cortices.
All of the subjects considered here found the experience of musical hallucinations distressing and requested treatment. Only one subject benefited from any treatment, in the form of improved amplification. I recommend that subjects within this group are assessed by an audiology service to decide if improved amplification may be of benefit, but in general the prognosis for improving the condition is poor.
Subject details
| Subject | Age | Sex | Hand* | Musical† | Index event | Vascular disease | Symptomatic deafness | Measured deafness‡ (dB ISO) | Tinnitus | Vertigo | Lateralizing neurological signs | Structural imaging | EEG |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| *Handedness on six-point Annett scale (Annett, 1970); †defined as passing a grade examination in any musical instrument; ‡mean hearing level (dB ISO) for the two ears measured at intervals of 1 octave from 250 Hz to 8 kHz. HT, hypertension. | |||||||||||||
| 1 | 73 | M | 6/6 R | No | No | Stable angina | <5 yr bilateral | 51 | No | No | No | MRI normal | Normal |
| 2 | 71 | M | 6/6 R | No | No | HT | Noise damage 1950s (tank crew) then progressive bilateral loss | 53 | No | Occasional with head movement | No | MRI signal change in white matter cerebral hemispheres | |
| 3 | 78 | F | 6/6 R | No | No | HT | Progressive bilateral loss >40 yr | 95 | No | No | No | MRI normal | Normal |
| 4 | 58 | F | 6/6 R | No | No | No risk | 23 yr progressive bilateral loss | 82 | Yes | Yes during early deafness | No | MRI normal | No specific abnormality |
| 5 | 65 | M | 6/6 R | No | Head injuries aged 15, 57 63 yr | No risk factors | L deafness after after head injury at age 15 yr R deafness after head injury aged 57 yr, then bilateral progression | 105 | Yes | No | No | MRI signal change in hemispheric white matter/pons | Minor slowing |
| 6 | 82 | F | 6/6 R | No | Posterior circulation stroke aged 79 yr | Single stroke without risk factors | Progressive bilateral,15 yr | 69 | No | No | No | R occipital lobe cyst on CT | Normal |
| Subject | Age | Sex | Hand* | Musical† | Index event | Vascular disease | Symptomatic deafness | Measured deafness‡ (dB ISO) | Tinnitus | Vertigo | Lateralizing neurological signs | Structural imaging | EEG |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| *Handedness on six-point Annett scale (Annett, 1970); †defined as passing a grade examination in any musical instrument; ‡mean hearing level (dB ISO) for the two ears measured at intervals of 1 octave from 250 Hz to 8 kHz. HT, hypertension. | |||||||||||||
| 1 | 73 | M | 6/6 R | No | No | Stable angina | <5 yr bilateral | 51 | No | No | No | MRI normal | Normal |
| 2 | 71 | M | 6/6 R | No | No | HT | Noise damage 1950s (tank crew) then progressive bilateral loss | 53 | No | Occasional with head movement | No | MRI signal change in white matter cerebral hemispheres | |
| 3 | 78 | F | 6/6 R | No | No | HT | Progressive bilateral loss >40 yr | 95 | No | No | No | MRI normal | Normal |
| 4 | 58 | F | 6/6 R | No | No | No risk | 23 yr progressive bilateral loss | 82 | Yes | Yes during early deafness | No | MRI normal | No specific abnormality |
| 5 | 65 | M | 6/6 R | No | Head injuries aged 15, 57 63 yr | No risk factors | L deafness after after head injury at age 15 yr R deafness after head injury aged 57 yr, then bilateral progression | 105 | Yes | No | No | MRI signal change in hemispheric white matter/pons | Minor slowing |
| 6 | 82 | F | 6/6 R | No | Posterior circulation stroke aged 79 yr | Single stroke without risk factors | Progressive bilateral,15 yr | 69 | No | No | No | R occipital lobe cyst on CT | Normal |
Subject details
| Subject | Age | Sex | Hand* | Musical† | Index event | Vascular disease | Symptomatic deafness | Measured deafness‡ (dB ISO) | Tinnitus | Vertigo | Lateralizing neurological signs | Structural imaging | EEG |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| *Handedness on six-point Annett scale (Annett, 1970); †defined as passing a grade examination in any musical instrument; ‡mean hearing level (dB ISO) for the two ears measured at intervals of 1 octave from 250 Hz to 8 kHz. HT, hypertension. | |||||||||||||
| 1 | 73 | M | 6/6 R | No | No | Stable angina | <5 yr bilateral | 51 | No | No | No | MRI normal | Normal |
| 2 | 71 | M | 6/6 R | No | No | HT | Noise damage 1950s (tank crew) then progressive bilateral loss | 53 | No | Occasional with head movement | No | MRI signal change in white matter cerebral hemispheres | |
| 3 | 78 | F | 6/6 R | No | No | HT | Progressive bilateral loss >40 yr | 95 | No | No | No | MRI normal | Normal |
| 4 | 58 | F | 6/6 R | No | No | No risk | 23 yr progressive bilateral loss | 82 | Yes | Yes during early deafness | No | MRI normal | No specific abnormality |
| 5 | 65 | M | 6/6 R | No | Head injuries aged 15, 57 63 yr | No risk factors | L deafness after after head injury at age 15 yr R deafness after head injury aged 57 yr, then bilateral progression | 105 | Yes | No | No | MRI signal change in hemispheric white matter/pons | Minor slowing |
| 6 | 82 | F | 6/6 R | No | Posterior circulation stroke aged 79 yr | Single stroke without risk factors | Progressive bilateral,15 yr | 69 | No | No | No | R occipital lobe cyst on CT | Normal |
| Subject | Age | Sex | Hand* | Musical† | Index event | Vascular disease | Symptomatic deafness | Measured deafness‡ (dB ISO) | Tinnitus | Vertigo | Lateralizing neurological signs | Structural imaging | EEG |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| *Handedness on six-point Annett scale (Annett, 1970); †defined as passing a grade examination in any musical instrument; ‡mean hearing level (dB ISO) for the two ears measured at intervals of 1 octave from 250 Hz to 8 kHz. HT, hypertension. | |||||||||||||
| 1 | 73 | M | 6/6 R | No | No | Stable angina | <5 yr bilateral | 51 | No | No | No | MRI normal | Normal |
| 2 | 71 | M | 6/6 R | No | No | HT | Noise damage 1950s (tank crew) then progressive bilateral loss | 53 | No | Occasional with head movement | No | MRI signal change in white matter cerebral hemispheres | |
| 3 | 78 | F | 6/6 R | No | No | HT | Progressive bilateral loss >40 yr | 95 | No | No | No | MRI normal | Normal |
| 4 | 58 | F | 6/6 R | No | No | No risk | 23 yr progressive bilateral loss | 82 | Yes | Yes during early deafness | No | MRI normal | No specific abnormality |
| 5 | 65 | M | 6/6 R | No | Head injuries aged 15, 57 63 yr | No risk factors | L deafness after after head injury at age 15 yr R deafness after head injury aged 57 yr, then bilateral progression | 105 | Yes | No | No | MRI signal change in hemispheric white matter/pons | Minor slowing |
| 6 | 82 | F | 6/6 R | No | Posterior circulation stroke aged 79 yr | Single stroke without risk factors | Progressive bilateral,15 yr | 69 | No | No | No | R occipital lobe cyst on CT | Normal |
Phenomenology
| Scan | Score | Tune/artist | Instrument | Singer | Distinct lyrics |
|---|---|---|---|---|---|
| Subject 1 | |||||
| 1 | 3 | `Football's coming home' | No | Several male | No |
| 2 | 3 | `Football's coming home' | No | Several male | No |
| 3 | 2 | `Football's coming home' | No | Several male | No |
| 4 | 2 | `Football's coming home' | No | Several male | No |
| 5 | 2 | `Football's coming home' | No | Several male | No |
| 6 | 2 | `Football's coming home' | No | Several male | No |
| 7 | 4 | `Football's coming home' | No | Several male | No |
| 8 | 4 | `Football's coming home' | No | Several male | No |
| 9 | 4 | `Football's coming home' | No | Several male | No |
| 10 | 5 | `Football's coming home' | No | Several male | No |
| 11 | 5 | `Football's coming home' | No | Several male | No |
| 12 | 4 | `Football's coming home' | No | Several male | No |
| Subject 2 | |||||
| 1 | 7 | `What can I do?', Corrs | No | Several female | Yes |
| 2 | 7 | `24 hours from Tulsa' | No | One male | Yes |
| 3 | 7 | `I'll take you home, Kathleen' | No | Single male | Yes |
| 4 | 7 | `You are my heart's delight' | Piano | Single male | Yes |
| 5 | 7 | `Edelweiss' | Orchestra | 2 female | Yes |
| 6 | 7 | `Maria' | Guitar | 1 male | Yes |
| 7 | 1 | No music | No | No | No |
| 8 | 2 | Unrecognized tune | Orchestra | No | No |
| 9 | 1 | No music | No | No | No |
| 10 | 1 | No music | No | No | No |
| 11 | 4 | `Sweatheart' | No | 1 male/1 female | Yes |
| 12 | 1 | No music | No | No | No |
| Subject 3 | |||||
| 1 | 5 | `While shepherds' | Organ | 1 male/1 female | Yes |
| 2 | 4 | `While shepherds' | Organ | 1 male/1 female | Yes |
| 3 | 6 | `Poor old Joe' | Organ | 1 male | Yes |
| 4 | 7 | `While shepherds' | Organ | No | No |
| 5 | 6 | `All things bright' | Organ | No | No |
| 6 | 7 | `All things bright' | Organ | Several | Yes |
| 7 | 7 | `Daisy, daisy' | Organ | Single male | Yes |
| 8 | 7 | `Daisy, daisy' | Organ | Single male | Yes |
| 9 | 7 | `Stand up for Jesus' | Organ | Single male | Yes |
| 10 | 7 | `Auld Lang Syne' | Organ | 1 male/1 female | Yes |
| 11 | 7 | `Auld Lang Syne' | Organ | 1 male/1 female | Yes |
| 12 | 7 | `Daisy, daisy' | Organ | 1 male/1 female | Yes |
| Subject 4 | |||||
| 1 | 2 | `Oh my beloved father' | Buzzy pitch | No | No |
| 2 | 3 | French Christmas carol | Buzzy pitch | No | No |
| 3 | 3 | French Christmas carol | Buzzy pitch | No | No |
| 4 | 2 | `Sweet Caroline', Diamond | Buzzy pitch | No | No |
| 5 | 3 | `Pomp and circumstance' | Buzzy pitch | No | No |
| 6 | 4 | The trout | Buzzy pitch | No | No |
| 7 | 3 | `Sound of music' medley | Buzzy pitch | No | No |
| 8 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 9 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 10 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 11 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 12 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| Subject 5 | |||||
| 1 | 5 | `Che sera sera' | Band | Yes several | Yes |
| 2 | 5 | `Lilly Marlene' | Band | Yes several | Yes |
| 3 | 5 | `Hey there' | Band | Yes several | Yes |
| 4 | 5 | `Runaway train' | Band | Yes several | Yes |
| 5 | 5 | `Three caballros' | Band | Yes several | Yes |
| Subject 5 | |||||
| 6 | 5 | `Danny boy' | Band | Yes several | Yes |
| 7 | 5 | `When you're in love' | Band | Yes, 4 | Yes |
| 8 | 5 | Irish national anthem | Band | Yes, large group | Yes |
| 9 | 5 | `More than ever' | Band | Yes | Yes |
| 10 | 5 | `Yellow rose of Texas' | Band | Yes | Yes |
| 11 | 5 | `Lilly Marlene' | Band | Yes | Yes |
| 12 | 5 | `Lilly Marlene' | Band | Yes | Yes |
| Subject 6 | |||||
| 1 | 3 | `Don't laugh at me 'cause I'm a fool' | Band | Yes, several | Yes |
| 2 | 5 | `Moonlight and roses' | Band | Yes, several | Yes |
| 3 | 5 | `Dreaming' | Piano | Yes, several | Yes |
| 4 | 5 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 5 | 6 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 6 | 6 | `Pal of my cradle days' | Piano | Yes, several | Yes |
| 7 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 8 | 7 | `Don't laugh at me `cause I'm a fool' | Piano | Yes, several | Yes |
| 9 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 10 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 11 | 7 | `Good night Vienna' | Piano | Yes, several | Yes |
| 12 | 7 | `Good night Vienna' | Band | Yes, several | Yes |
| Scan | Score | Tune/artist | Instrument | Singer | Distinct lyrics |
|---|---|---|---|---|---|
| Subject 1 | |||||
| 1 | 3 | `Football's coming home' | No | Several male | No |
| 2 | 3 | `Football's coming home' | No | Several male | No |
| 3 | 2 | `Football's coming home' | No | Several male | No |
| 4 | 2 | `Football's coming home' | No | Several male | No |
| 5 | 2 | `Football's coming home' | No | Several male | No |
| 6 | 2 | `Football's coming home' | No | Several male | No |
| 7 | 4 | `Football's coming home' | No | Several male | No |
| 8 | 4 | `Football's coming home' | No | Several male | No |
| 9 | 4 | `Football's coming home' | No | Several male | No |
| 10 | 5 | `Football's coming home' | No | Several male | No |
| 11 | 5 | `Football's coming home' | No | Several male | No |
| 12 | 4 | `Football's coming home' | No | Several male | No |
| Subject 2 | |||||
| 1 | 7 | `What can I do?', Corrs | No | Several female | Yes |
| 2 | 7 | `24 hours from Tulsa' | No | One male | Yes |
| 3 | 7 | `I'll take you home, Kathleen' | No | Single male | Yes |
| 4 | 7 | `You are my heart's delight' | Piano | Single male | Yes |
| 5 | 7 | `Edelweiss' | Orchestra | 2 female | Yes |
| 6 | 7 | `Maria' | Guitar | 1 male | Yes |
| 7 | 1 | No music | No | No | No |
| 8 | 2 | Unrecognized tune | Orchestra | No | No |
| 9 | 1 | No music | No | No | No |
| 10 | 1 | No music | No | No | No |
| 11 | 4 | `Sweatheart' | No | 1 male/1 female | Yes |
| 12 | 1 | No music | No | No | No |
| Subject 3 | |||||
| 1 | 5 | `While shepherds' | Organ | 1 male/1 female | Yes |
| 2 | 4 | `While shepherds' | Organ | 1 male/1 female | Yes |
| 3 | 6 | `Poor old Joe' | Organ | 1 male | Yes |
| 4 | 7 | `While shepherds' | Organ | No | No |
| 5 | 6 | `All things bright' | Organ | No | No |
| 6 | 7 | `All things bright' | Organ | Several | Yes |
| 7 | 7 | `Daisy, daisy' | Organ | Single male | Yes |
| 8 | 7 | `Daisy, daisy' | Organ | Single male | Yes |
| 9 | 7 | `Stand up for Jesus' | Organ | Single male | Yes |
| 10 | 7 | `Auld Lang Syne' | Organ | 1 male/1 female | Yes |
| 11 | 7 | `Auld Lang Syne' | Organ | 1 male/1 female | Yes |
| 12 | 7 | `Daisy, daisy' | Organ | 1 male/1 female | Yes |
| Subject 4 | |||||
| 1 | 2 | `Oh my beloved father' | Buzzy pitch | No | No |
| 2 | 3 | French Christmas carol | Buzzy pitch | No | No |
| 3 | 3 | French Christmas carol | Buzzy pitch | No | No |
| 4 | 2 | `Sweet Caroline', Diamond | Buzzy pitch | No | No |
| 5 | 3 | `Pomp and circumstance' | Buzzy pitch | No | No |
| 6 | 4 | The trout | Buzzy pitch | No | No |
| 7 | 3 | `Sound of music' medley | Buzzy pitch | No | No |
| 8 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 9 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 10 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 11 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 12 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| Subject 5 | |||||
| 1 | 5 | `Che sera sera' | Band | Yes several | Yes |
| 2 | 5 | `Lilly Marlene' | Band | Yes several | Yes |
| 3 | 5 | `Hey there' | Band | Yes several | Yes |
| 4 | 5 | `Runaway train' | Band | Yes several | Yes |
| 5 | 5 | `Three caballros' | Band | Yes several | Yes |
| Subject 5 | |||||
| 6 | 5 | `Danny boy' | Band | Yes several | Yes |
| 7 | 5 | `When you're in love' | Band | Yes, 4 | Yes |
| 8 | 5 | Irish national anthem | Band | Yes, large group | Yes |
| 9 | 5 | `More than ever' | Band | Yes | Yes |
| 10 | 5 | `Yellow rose of Texas' | Band | Yes | Yes |
| 11 | 5 | `Lilly Marlene' | Band | Yes | Yes |
| 12 | 5 | `Lilly Marlene' | Band | Yes | Yes |
| Subject 6 | |||||
| 1 | 3 | `Don't laugh at me 'cause I'm a fool' | Band | Yes, several | Yes |
| 2 | 5 | `Moonlight and roses' | Band | Yes, several | Yes |
| 3 | 5 | `Dreaming' | Piano | Yes, several | Yes |
| 4 | 5 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 5 | 6 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 6 | 6 | `Pal of my cradle days' | Piano | Yes, several | Yes |
| 7 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 8 | 7 | `Don't laugh at me `cause I'm a fool' | Piano | Yes, several | Yes |
| 9 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 10 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 11 | 7 | `Good night Vienna' | Piano | Yes, several | Yes |
| 12 | 7 | `Good night Vienna' | Band | Yes, several | Yes |
Phenomenology
| Scan | Score | Tune/artist | Instrument | Singer | Distinct lyrics |
|---|---|---|---|---|---|
| Subject 1 | |||||
| 1 | 3 | `Football's coming home' | No | Several male | No |
| 2 | 3 | `Football's coming home' | No | Several male | No |
| 3 | 2 | `Football's coming home' | No | Several male | No |
| 4 | 2 | `Football's coming home' | No | Several male | No |
| 5 | 2 | `Football's coming home' | No | Several male | No |
| 6 | 2 | `Football's coming home' | No | Several male | No |
| 7 | 4 | `Football's coming home' | No | Several male | No |
| 8 | 4 | `Football's coming home' | No | Several male | No |
| 9 | 4 | `Football's coming home' | No | Several male | No |
| 10 | 5 | `Football's coming home' | No | Several male | No |
| 11 | 5 | `Football's coming home' | No | Several male | No |
| 12 | 4 | `Football's coming home' | No | Several male | No |
| Subject 2 | |||||
| 1 | 7 | `What can I do?', Corrs | No | Several female | Yes |
| 2 | 7 | `24 hours from Tulsa' | No | One male | Yes |
| 3 | 7 | `I'll take you home, Kathleen' | No | Single male | Yes |
| 4 | 7 | `You are my heart's delight' | Piano | Single male | Yes |
| 5 | 7 | `Edelweiss' | Orchestra | 2 female | Yes |
| 6 | 7 | `Maria' | Guitar | 1 male | Yes |
| 7 | 1 | No music | No | No | No |
| 8 | 2 | Unrecognized tune | Orchestra | No | No |
| 9 | 1 | No music | No | No | No |
| 10 | 1 | No music | No | No | No |
| 11 | 4 | `Sweatheart' | No | 1 male/1 female | Yes |
| 12 | 1 | No music | No | No | No |
| Subject 3 | |||||
| 1 | 5 | `While shepherds' | Organ | 1 male/1 female | Yes |
| 2 | 4 | `While shepherds' | Organ | 1 male/1 female | Yes |
| 3 | 6 | `Poor old Joe' | Organ | 1 male | Yes |
| 4 | 7 | `While shepherds' | Organ | No | No |
| 5 | 6 | `All things bright' | Organ | No | No |
| 6 | 7 | `All things bright' | Organ | Several | Yes |
| 7 | 7 | `Daisy, daisy' | Organ | Single male | Yes |
| 8 | 7 | `Daisy, daisy' | Organ | Single male | Yes |
| 9 | 7 | `Stand up for Jesus' | Organ | Single male | Yes |
| 10 | 7 | `Auld Lang Syne' | Organ | 1 male/1 female | Yes |
| 11 | 7 | `Auld Lang Syne' | Organ | 1 male/1 female | Yes |
| 12 | 7 | `Daisy, daisy' | Organ | 1 male/1 female | Yes |
| Subject 4 | |||||
| 1 | 2 | `Oh my beloved father' | Buzzy pitch | No | No |
| 2 | 3 | French Christmas carol | Buzzy pitch | No | No |
| 3 | 3 | French Christmas carol | Buzzy pitch | No | No |
| 4 | 2 | `Sweet Caroline', Diamond | Buzzy pitch | No | No |
| 5 | 3 | `Pomp and circumstance' | Buzzy pitch | No | No |
| 6 | 4 | The trout | Buzzy pitch | No | No |
| 7 | 3 | `Sound of music' medley | Buzzy pitch | No | No |
| 8 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 9 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 10 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 11 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 12 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| Subject 5 | |||||
| 1 | 5 | `Che sera sera' | Band | Yes several | Yes |
| 2 | 5 | `Lilly Marlene' | Band | Yes several | Yes |
| 3 | 5 | `Hey there' | Band | Yes several | Yes |
| 4 | 5 | `Runaway train' | Band | Yes several | Yes |
| 5 | 5 | `Three caballros' | Band | Yes several | Yes |
| Subject 5 | |||||
| 6 | 5 | `Danny boy' | Band | Yes several | Yes |
| 7 | 5 | `When you're in love' | Band | Yes, 4 | Yes |
| 8 | 5 | Irish national anthem | Band | Yes, large group | Yes |
| 9 | 5 | `More than ever' | Band | Yes | Yes |
| 10 | 5 | `Yellow rose of Texas' | Band | Yes | Yes |
| 11 | 5 | `Lilly Marlene' | Band | Yes | Yes |
| 12 | 5 | `Lilly Marlene' | Band | Yes | Yes |
| Subject 6 | |||||
| 1 | 3 | `Don't laugh at me 'cause I'm a fool' | Band | Yes, several | Yes |
| 2 | 5 | `Moonlight and roses' | Band | Yes, several | Yes |
| 3 | 5 | `Dreaming' | Piano | Yes, several | Yes |
| 4 | 5 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 5 | 6 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 6 | 6 | `Pal of my cradle days' | Piano | Yes, several | Yes |
| 7 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 8 | 7 | `Don't laugh at me `cause I'm a fool' | Piano | Yes, several | Yes |
| 9 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 10 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 11 | 7 | `Good night Vienna' | Piano | Yes, several | Yes |
| 12 | 7 | `Good night Vienna' | Band | Yes, several | Yes |
| Scan | Score | Tune/artist | Instrument | Singer | Distinct lyrics |
|---|---|---|---|---|---|
| Subject 1 | |||||
| 1 | 3 | `Football's coming home' | No | Several male | No |
| 2 | 3 | `Football's coming home' | No | Several male | No |
| 3 | 2 | `Football's coming home' | No | Several male | No |
| 4 | 2 | `Football's coming home' | No | Several male | No |
| 5 | 2 | `Football's coming home' | No | Several male | No |
| 6 | 2 | `Football's coming home' | No | Several male | No |
| 7 | 4 | `Football's coming home' | No | Several male | No |
| 8 | 4 | `Football's coming home' | No | Several male | No |
| 9 | 4 | `Football's coming home' | No | Several male | No |
| 10 | 5 | `Football's coming home' | No | Several male | No |
| 11 | 5 | `Football's coming home' | No | Several male | No |
| 12 | 4 | `Football's coming home' | No | Several male | No |
| Subject 2 | |||||
| 1 | 7 | `What can I do?', Corrs | No | Several female | Yes |
| 2 | 7 | `24 hours from Tulsa' | No | One male | Yes |
| 3 | 7 | `I'll take you home, Kathleen' | No | Single male | Yes |
| 4 | 7 | `You are my heart's delight' | Piano | Single male | Yes |
| 5 | 7 | `Edelweiss' | Orchestra | 2 female | Yes |
| 6 | 7 | `Maria' | Guitar | 1 male | Yes |
| 7 | 1 | No music | No | No | No |
| 8 | 2 | Unrecognized tune | Orchestra | No | No |
| 9 | 1 | No music | No | No | No |
| 10 | 1 | No music | No | No | No |
| 11 | 4 | `Sweatheart' | No | 1 male/1 female | Yes |
| 12 | 1 | No music | No | No | No |
| Subject 3 | |||||
| 1 | 5 | `While shepherds' | Organ | 1 male/1 female | Yes |
| 2 | 4 | `While shepherds' | Organ | 1 male/1 female | Yes |
| 3 | 6 | `Poor old Joe' | Organ | 1 male | Yes |
| 4 | 7 | `While shepherds' | Organ | No | No |
| 5 | 6 | `All things bright' | Organ | No | No |
| 6 | 7 | `All things bright' | Organ | Several | Yes |
| 7 | 7 | `Daisy, daisy' | Organ | Single male | Yes |
| 8 | 7 | `Daisy, daisy' | Organ | Single male | Yes |
| 9 | 7 | `Stand up for Jesus' | Organ | Single male | Yes |
| 10 | 7 | `Auld Lang Syne' | Organ | 1 male/1 female | Yes |
| 11 | 7 | `Auld Lang Syne' | Organ | 1 male/1 female | Yes |
| 12 | 7 | `Daisy, daisy' | Organ | 1 male/1 female | Yes |
| Subject 4 | |||||
| 1 | 2 | `Oh my beloved father' | Buzzy pitch | No | No |
| 2 | 3 | French Christmas carol | Buzzy pitch | No | No |
| 3 | 3 | French Christmas carol | Buzzy pitch | No | No |
| 4 | 2 | `Sweet Caroline', Diamond | Buzzy pitch | No | No |
| 5 | 3 | `Pomp and circumstance' | Buzzy pitch | No | No |
| 6 | 4 | The trout | Buzzy pitch | No | No |
| 7 | 3 | `Sound of music' medley | Buzzy pitch | No | No |
| 8 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 9 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 10 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 11 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| 12 | 5 | `Doe a deer' | Buzzy pitch | No | No |
| Subject 5 | |||||
| 1 | 5 | `Che sera sera' | Band | Yes several | Yes |
| 2 | 5 | `Lilly Marlene' | Band | Yes several | Yes |
| 3 | 5 | `Hey there' | Band | Yes several | Yes |
| 4 | 5 | `Runaway train' | Band | Yes several | Yes |
| 5 | 5 | `Three caballros' | Band | Yes several | Yes |
| Subject 5 | |||||
| 6 | 5 | `Danny boy' | Band | Yes several | Yes |
| 7 | 5 | `When you're in love' | Band | Yes, 4 | Yes |
| 8 | 5 | Irish national anthem | Band | Yes, large group | Yes |
| 9 | 5 | `More than ever' | Band | Yes | Yes |
| 10 | 5 | `Yellow rose of Texas' | Band | Yes | Yes |
| 11 | 5 | `Lilly Marlene' | Band | Yes | Yes |
| 12 | 5 | `Lilly Marlene' | Band | Yes | Yes |
| Subject 6 | |||||
| 1 | 3 | `Don't laugh at me 'cause I'm a fool' | Band | Yes, several | Yes |
| 2 | 5 | `Moonlight and roses' | Band | Yes, several | Yes |
| 3 | 5 | `Dreaming' | Piano | Yes, several | Yes |
| 4 | 5 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 5 | 6 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 6 | 6 | `Pal of my cradle days' | Piano | Yes, several | Yes |
| 7 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 8 | 7 | `Don't laugh at me `cause I'm a fool' | Piano | Yes, several | Yes |
| 9 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 10 | 7 | `Just a rose in a garden of weeds' | Band | Yes, several | Yes |
| 11 | 7 | `Good night Vienna' | Piano | Yes, several | Yes |
| 12 | 7 | `Good night Vienna' | Band | Yes, several | Yes |
Functional imaging (areas of significant activation as a linear function of hallucinosis intensity): group analysis for Subjects 1–4
| Region | Coordinates (mm) | Z score | ||
|---|---|---|---|---|
| x | y | z | ||
| Clusters demonstrating significant activation as a linear function of hallucinosis intensity at the P < 0.05 level (corrected for multiple comparisons using Gaussian field theory) are shown. The three most significant maxima within each cluster are shown, except for the large cluster involving the right basal ganglia and frontal cortex, where six local maxima are shown. | ||||
| Right basal ganglia (putamen/globus pallidus) | 24 | –10 | 2 | 5.45 |
| 28 | –8 | –6 | 4.77 | |
| 16 | –10 | 0 | 4.55 | |
| Right frontal | 54 | 12 | –4 | 4.15 |
| 56 | 12 | 0 | 4.09 | |
| 36 | 8 | –6 | 4.07 | |
| Left sylvian fissure | –40 | –24 | 24 | 5.24 |
| –34 | –14 | 24 | 4.77 | |
| –34 | –20 | 14 | 3.69 | |
| Right posterior temporal | 72 | –38 | –16 | 5.07 |
| 74 | –38 | –8 | 4.62 | |
| Left cerebellum | –42 | –82 | –24 | 4.66 |
| –44 | –82 | –36 | 4.31 | |
| –32 | –78 | –28 | 4.23 | |
| Right cerebellum | 16 | –54 | –32 | 4.43 |
| 22 | –62 | –26 | 3.65 | |
| 24 | –70 | –22 | 3.44 | |
| Left frontal lobe | –32 | 16 | 34 | 4.25 |
| –50 | 28 | 32 | 3.71 | |
| –38 | 20 | 40 | 3.70 | |
| –20 | 36 | 2 | 4.23 | |
| Left posterior temporal lobe | –62 | –38 | 4 | 4.02 |
| –70 | –44 | 8 | 3.88 | |
| –60 | –30 | 0 | 3.69 | |
| Region | Coordinates (mm) | Z score | ||
|---|---|---|---|---|
| x | y | z | ||
| Clusters demonstrating significant activation as a linear function of hallucinosis intensity at the P < 0.05 level (corrected for multiple comparisons using Gaussian field theory) are shown. The three most significant maxima within each cluster are shown, except for the large cluster involving the right basal ganglia and frontal cortex, where six local maxima are shown. | ||||
| Right basal ganglia (putamen/globus pallidus) | 24 | –10 | 2 | 5.45 |
| 28 | –8 | –6 | 4.77 | |
| 16 | –10 | 0 | 4.55 | |
| Right frontal | 54 | 12 | –4 | 4.15 |
| 56 | 12 | 0 | 4.09 | |
| 36 | 8 | –6 | 4.07 | |
| Left sylvian fissure | –40 | –24 | 24 | 5.24 |
| –34 | –14 | 24 | 4.77 | |
| –34 | –20 | 14 | 3.69 | |
| Right posterior temporal | 72 | –38 | –16 | 5.07 |
| 74 | –38 | –8 | 4.62 | |
| Left cerebellum | –42 | –82 | –24 | 4.66 |
| –44 | –82 | –36 | 4.31 | |
| –32 | –78 | –28 | 4.23 | |
| Right cerebellum | 16 | –54 | –32 | 4.43 |
| 22 | –62 | –26 | 3.65 | |
| 24 | –70 | –22 | 3.44 | |
| Left frontal lobe | –32 | 16 | 34 | 4.25 |
| –50 | 28 | 32 | 3.71 | |
| –38 | 20 | 40 | 3.70 | |
| –20 | 36 | 2 | 4.23 | |
| Left posterior temporal lobe | –62 | –38 | 4 | 4.02 |
| –70 | –44 | 8 | 3.88 | |
| –60 | –30 | 0 | 3.69 | |
Functional imaging (areas of significant activation as a linear function of hallucinosis intensity): group analysis for Subjects 1–4
| Region | Coordinates (mm) | Z score | ||
|---|---|---|---|---|
| x | y | z | ||
| Clusters demonstrating significant activation as a linear function of hallucinosis intensity at the P < 0.05 level (corrected for multiple comparisons using Gaussian field theory) are shown. The three most significant maxima within each cluster are shown, except for the large cluster involving the right basal ganglia and frontal cortex, where six local maxima are shown. | ||||
| Right basal ganglia (putamen/globus pallidus) | 24 | –10 | 2 | 5.45 |
| 28 | –8 | –6 | 4.77 | |
| 16 | –10 | 0 | 4.55 | |
| Right frontal | 54 | 12 | –4 | 4.15 |
| 56 | 12 | 0 | 4.09 | |
| 36 | 8 | –6 | 4.07 | |
| Left sylvian fissure | –40 | –24 | 24 | 5.24 |
| –34 | –14 | 24 | 4.77 | |
| –34 | –20 | 14 | 3.69 | |
| Right posterior temporal | 72 | –38 | –16 | 5.07 |
| 74 | –38 | –8 | 4.62 | |
| Left cerebellum | –42 | –82 | –24 | 4.66 |
| –44 | –82 | –36 | 4.31 | |
| –32 | –78 | –28 | 4.23 | |
| Right cerebellum | 16 | –54 | –32 | 4.43 |
| 22 | –62 | –26 | 3.65 | |
| 24 | –70 | –22 | 3.44 | |
| Left frontal lobe | –32 | 16 | 34 | 4.25 |
| –50 | 28 | 32 | 3.71 | |
| –38 | 20 | 40 | 3.70 | |
| –20 | 36 | 2 | 4.23 | |
| Left posterior temporal lobe | –62 | –38 | 4 | 4.02 |
| –70 | –44 | 8 | 3.88 | |
| –60 | –30 | 0 | 3.69 | |
| Region | Coordinates (mm) | Z score | ||
|---|---|---|---|---|
| x | y | z | ||
| Clusters demonstrating significant activation as a linear function of hallucinosis intensity at the P < 0.05 level (corrected for multiple comparisons using Gaussian field theory) are shown. The three most significant maxima within each cluster are shown, except for the large cluster involving the right basal ganglia and frontal cortex, where six local maxima are shown. | ||||
| Right basal ganglia (putamen/globus pallidus) | 24 | –10 | 2 | 5.45 |
| 28 | –8 | –6 | 4.77 | |
| 16 | –10 | 0 | 4.55 | |
| Right frontal | 54 | 12 | –4 | 4.15 |
| 56 | 12 | 0 | 4.09 | |
| 36 | 8 | –6 | 4.07 | |
| Left sylvian fissure | –40 | –24 | 24 | 5.24 |
| –34 | –14 | 24 | 4.77 | |
| –34 | –20 | 14 | 3.69 | |
| Right posterior temporal | 72 | –38 | –16 | 5.07 |
| 74 | –38 | –8 | 4.62 | |
| Left cerebellum | –42 | –82 | –24 | 4.66 |
| –44 | –82 | –36 | 4.31 | |
| –32 | –78 | –28 | 4.23 | |
| Right cerebellum | 16 | –54 | –32 | 4.43 |
| 22 | –62 | –26 | 3.65 | |
| 24 | –70 | –22 | 3.44 | |
| Left frontal lobe | –32 | 16 | 34 | 4.25 |
| –50 | 28 | 32 | 3.71 | |
| –38 | 20 | 40 | 3.70 | |
| –20 | 36 | 2 | 4.23 | |
| Left posterior temporal lobe | –62 | –38 | 4 | 4.02 |
| –70 | –44 | 8 | 3.88 | |
| –60 | –30 | 0 | 3.69 | |
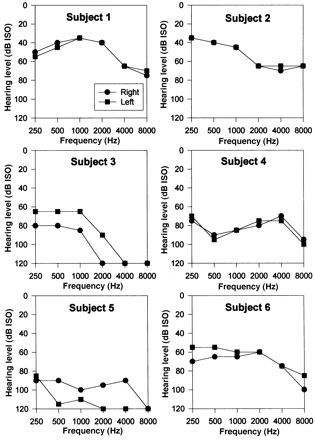
Pure-tone audiograms.
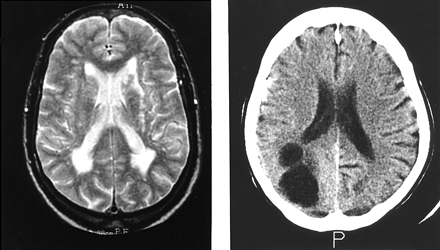
Structural imaging of Subjects 2 and 6. MRI of Subject 2 (left) shows multiple vascular lesions in the hemispheric deep white matter. CT scan of Subject 6 (right) shows a large arachnoid cyst in the right occipital lobe.
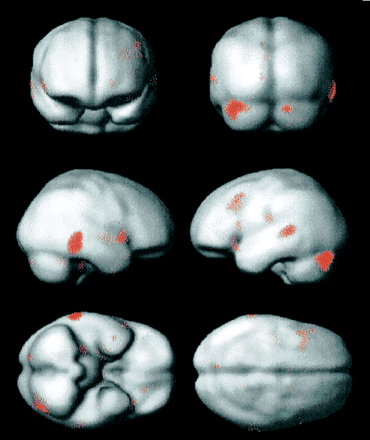
Functional imaging (correlated activity with hallucination strength surface reading). PET group analysis for Subjects 1–4 showing areas where regional cerebral blood flow increased as a linear function of hallucinosis intensity. The functional data shown were thresholded at the P < 0.001 (uncorrected) voxel level and rendered on to an MRI surface template with the same degree of smoothing as the PET data.
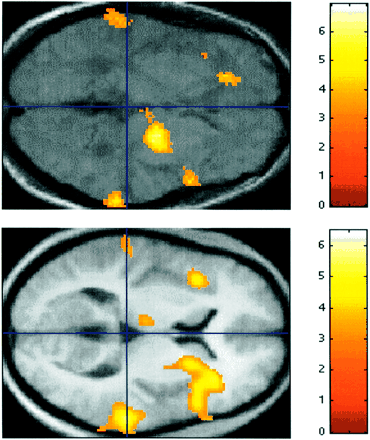
Functional imaging. (Top) PET group analysis for Subjects 1–4 showing areas where regional cerebral blood flow increased as a linear function of hallucinosis intensity (no sound stimulus). The data shown were rendered on to an axial section of the mean T1-weighted MRI for the four subjects. (Bottom) For comparison, data for the activation during the normal perception of patterned–segmented sound in nine normal subjects (Griffiths et al., 1999a) are shown superimposed on an axial section at the same level. Both sections are at the vertical level z = 0 mm. A line at y = –30 mm corresponds to the anteroposterior maximum for the planum temporale (Westbury et al., 1999). Notice the bilateral activation in the region of the planum temporale in both analyses.
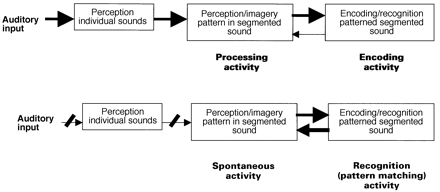
(Top) The processes occurring during the normal perception of pattern in segmented sound. (Bottom) The proposed basis for musical hallucinations, due to spontaneous activity in the module for the perception and imagery of pattern in segmented sound.
I wish to thank the patients, many of whom travelled long distances to undergo detailed assessment and scanning, the referring clinicians (R. Brenner, D. Burn, M. Jackson, J. Palace, S. Pereira, P. Reading, J. Spillane, B. Toone, R. Wise), members of the Neurootology Department, National Hospital for Neurology, Queen Square and the Department of Audiology, Kings College Hospital for carrying out the audiograms, the radiographers at the Wellcome Department of Cognitive Neurology for expert technical assistance, and R. S. J. Frackowiak for helpful comments. I am supported by the Wellcome Trust (UK).
References
Annett M. A classification of hand preference by association analysis.
Berrios GE. Musical hallucinations. A historical and clinical study.
Binder JR, Frost JA, Hammeke TA, Rao SM, Cox RW. Function of the left planum temporale in auditory and linguistic processing.
ffytche DH, Howard RJ, Brammer MJ, David A, Woodruff P, Williams S. The anatomy of conscious vision: an fMRI study of visual hallucinations.
Friston KJ, Ashburner J, Frith CD, Poline J-B, Heather JD, Frackowiak RSJ. Spatial registration and normalization of images.
Giraud AL, Chery-Croze S, Fischer G, Fischer C, Vighetto A, Gregoire M-C, et al. A selective imaging of tinnitus.
Griffiths TD, Jackson MC, Spillane JA, Friston KJ, Frackowiak RSJ. A neural substrate for musical hallucinosis.
Griffiths TD, Buechel C, Frackowiak RS, Patterson RD. Analysis of temporal structure in sound by the human brain.
Griffiths TD, Johnsrude I, Dean JL, Green GGR. A common neural substrate for the analysis of pitch and duration pattern in segmented sound?
Griffiths TD, Rees A, Green GGR. Disorders of human complex sound processing.
Halpern AR, Zatorre RJ. When that tune runs through your head: a PET investigation of auditory imagery for familiar melodies.
Inzelberg R, Vishnievskaya S, Korczyn AD. Transient musical hallucinosis [letter].
Keshavan MS, David AS, Steingard S, Lishman WA. Musical hallucinations: a review and synthesis.
Lockwood AH, Salvi RJ, Coad ML, Towsley ML, Wack DS, Murphy BW. The functional neuroanatomy of tinnitus: evidence for limbic system links and neural plasticity.
Mummery CJ, Ashburner J, Scott SK, Wise RJ. Functional neuroimaging of speech perception in six normal and two aphasic subjects.
Murata S, Naritomi H, Sawada T. Musical auditory hallucinations caused by a brainstem lesion.
Pantev C, Hoke M, Lutkenhoner B, Lehnertz K. Tonotopic organization of the auditory cortex: pitch versus frequency representation.
Paquier P, van Vugt P, Bal P, Cras P, Parizel PM, van Haesendonck J, et al. Transient musical hallucinosis of central origin: a review and clinical study.
Penhune VB, Zatorre RJ, MacDonald JD, Evans AC. Interhemispheric anatomical differences in human primary auditory cortex: probabilistic mapping and volume measurement from magnetic resonance scans.
Penhune VB, Zatorre RJ, Evans AC. Cerebellar contributions to motor timing: a PET study of auditory and visual rhythm reproduction.
Phillips DP, Hall SE. Response timing constraints on the cortical representation of sound time structure.
Rao SM, Harrington DL, Haaland KY, Bobholz JA, Cox RW, Binder JR. Distributed neural systems underlying the timing of movements.
Silbersweig DA, Stern E, Frith C, Cahill C, Holmes A, Grootoonk S, et al. A functional neuroanatomy of hallucinations in schizophrenia.
Tanabe H, Sawada T, Asai H, Okuda J, Shiraishi J. Lateralization phenomenon of complex auditory hallucinations.
Westbury CF, Zatorre RJ, Evans AC. Quantifying variability in the planum temporale: a probability map.
Zatorre RJ. Pitch perception of complex tones and human temporal-lobe function.
Zatorre RJ, Evans AC, Meyer E. Neural mechanisms underlying melodic perception and memory for pitch.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}